

Review
doi: 10.1105/tpc.001115.
Calmodulins and calcineurin B-like proteins: calcium sensors for specific signal response coupling in plants
Affiliations
- PMID: 12045290
- PMCID: PMC151268
- DOI: 10.1105/tpc.001115
Item in Clipboard
Review
Calmodulins and calcineurin B-like proteins: calcium sensors for specific signal response coupling in plants
Plant Cell.
2002.
No abstract available
Figures

Scheme of CaMs and CaM-Related Proteins in Plants. Typical CaMs (such as Arabidopsis [At] CaM2 and petunia [Ph] CaM81) contain four EF hands (red ovals). CaM-related proteins show additional domains (indicated by boxes) and/or either removal or addition of EF hands. For instance, Arabidopsis CaM11 contains an insertion of 22 Gln residues (22xQ) that follow the first four amino acids of the protein (Table 1). Petunia CaM53 and rice (Os) CaM61 contain a C-terminal extension that comprises a polybasic domain (+) and a CaaL box that specifies prenylation by GGTase-I (Rodriguez-Concepcion et al., 1999; Dong et al., 2002). A conserved C-terminal extension of unknown function is present in both Arabidopsis CaM10 and CaM12, but the latter contains two additional EF hands (Table 1). Extra EF hands and unrelated sequences also are found in the protein encoded by Arabidopsis TCH3, a touch-induced gene (Sistrunk et al., 1994). Substitution of a EF hand for a nonconserved sequence is observed in wheat (Ta) CaM2-1 (Yang et al., 1996) and tobacco (Nt) rgsCaM, which was found to be a suppressor of post-transcriptional gene silencing (Anandalakshmi et al., 2000).
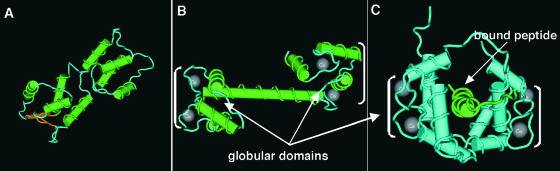
Structural Analysis of Apo-CaM, Ca2+-CaM, and the Ca2+-CaM–Target Complex. Strand-rod presentation of Apo-CaM (A) and Ca2+-CaM (B) showing substantial changes upon Ca2+ binding. (C) shows a solution structure solved by NMR of peptide-bound Ca2+-CaM. Peptide binding causes disruption of the flexible tether, bringing the globular domains closer to form a channel around the peptide. The majority of contacts between Ca2+-CaM and target peptide are nonspecific van der Waals bonds made by residues in the hydrophobic surfaces. Brackets indicate globular domains.

Subcellular and Tissue-Specific Localization of CaM53. (A) A CaM53-specific antibody (Rodriguez-Concepcion et al., 1999) was used for immunolocalization of the protein on a paraffin-embedded petunia longitudinal section showing a floral meristem. Secondary antibodies coupled to alkaline phosphatase were used to visualize the localization of cross-reacting polypeptides with bright-field illumination. (B) to (D) Subcellular localization was studied in petunia protoplasts transformed with the plasmid to express the full-length CaM53 protein fused to green fluorescent protein as described previously (Rodriguez-Concepcion et al., 1999). After electroporation, protoplasts were incubated for 16 hr in the presence of 5 μM mevinolin (B), 1 mM CaCl2
(C), or 1 mM EGTA (D). Localization of the green fluorescent protein–CaM53 fusion protein was detected by green fluorescence. Red fluorescence corresponds to chlorophyll. Arrow in (B) indicates the nucleus. (E) Ectopic overexpression of the wild-type CaM53 (CaM53wt) or a nonprenylated mutant version (CaM53mS) in Nicotiana benthamiana plants using the potato virus x viral vector induced distinct developmental abnormalities. An empty potato virus x vector–infected plant is shown as a control (Rodriguez-Concepcion et al., 1999). The photograph was taken 35 days after inoculation.
References
-
- Anandalakshmi, R., Marathe, R., Ge, X., Herr, J.M., Jr., Mau, C., Mallory, A., Pruss, G., Bowman, L., and Vance, V.B. (2000). A calmodulin-related protein that suppresses posttranscriptional gene silencing in plants. Science 290 142–144. - PubMed
-
- Antosiewicz, D.M., Polisensky, D.H., and Braam, J. (1995). Cellular localization of the Ca2+ binding TCH3 protein of Arabidopsis. Plant J. 8 623–636. - PubMed
-
- Arazi, T., Kaplan, B., and Fromm, H. (2000). A high-affinity calmodulin-binding site in a tobacco plasma-membrane channel protein coincides with a characteristic element of cyclic nucleotide-binding domains. Plant Mol. Biol. 42 591–601. - PubMed
-
- Arazi, T., Sunkar, R., Kaplan, B., and Fromm, H. (1999). A tobacco plasma membrane calmodulin-binding transporter confers Ni2+ tolerance and Pb2+ hypersensitivity in transgenic plants. Plant J. 20 171–182. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
