

Review
doi: 10.1105/tpc.002899.
Calcium at the crossroads of signaling
Collaborators,
Affiliations
- PMID: 12045291
- PMCID: PMC151269
- DOI: 10.1105/tpc.002899
Item in Clipboard
Review
Calcium at the crossroads of signaling
Plant Cell.
2002.
No abstract available
Figures
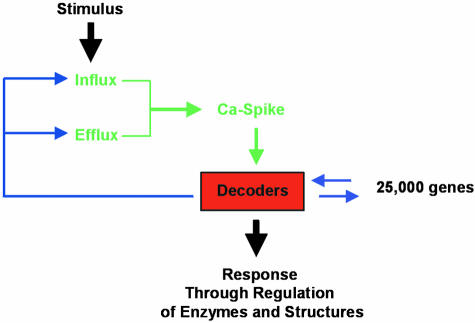
Decoding Calcium Signals Leads to a Specific Response at the Cellular Level. Various feedback mechanisms from the calcium sensor (or “decoder”) are possible. These could include the regulation of calcium spikes via the control of calcium permeable channel gating (e.g., through EF binding hands, or via Ca2+/CaM binding) or via control of pump activity.

Schematic Representation of Major Identified Ca2+ Transport Pathways in Arabidopsis Cell Membranes. Blue circles represent energized transport systems. ACA1, ACA4, ACA8 are autoinhibited calcium ATPases identified at a molecular level. The direction of Ca2+ pumping for ACA1 is hypothetical. ECA is an ER-type calcium ATPase. ACAx in the central vacuole and in the Golgi has not been identified at a molecular level. CAX1 is a Ca2+/H+ antiporter expected to be localized at the vacuolar membrane. Red squares represent Ca2+-permeable channels. At the plasma membrane, nonselective cation (NSC) channels, depolarization activated channels (DACs) and hyperpolarization activated channels (HACs) have been characterized at an electrophysiological but not at a molecular level. A two-pore channel (TPC1) has been shown to complement a yeast mutant deficient in Ca2+ uptake, but channel location is hypothetical. Using electrophysiolocal techniques, cyclic nucleotide gated channels (CNGC1 and CNGC2) were shown to be permeable to calcium. Plasma membrane location is again hypothetical. Glutamate receptors (GLRs) might be involved in the increase of cytosolic calcium concentrations and have been identified at a molecular level. The channels identified at endomembranes have been characterized at electrophysiological and biochemical but not molecular levels. InsP3R, putative Ins3P receptor; RyR, putative ryanodine receptor activated by cADPR; NAADP-activated channels also reside in the ER as shown. SV channel, slowly activating vacuolar channel; VVCa channel, vacuolar voltage-gated Ca2+ channel.

Topology Models of Putative Plasma Membrane Proteins Involved in Calcium Influx in the Cytosol in Arabidopsis. (A) The two-pore channel (TPC1) is composed of two EF calcium binding hands, which could be involved in the feedback control of the channel activity via cytosolic calcium concentration. The pore loop (P) is localized between the 5th and 6th transmembrane domains of each repeat. The 4th transmembrane domain in each repeat is enriched in basic residues, which might suggest that the channel is voltage gated. (B) CNGC structure also contains a P loop and, unlike counterparts in animals, overlapping of the calmodulin and cyclic nucleotide binding domains at the C terminus of the protein. (C) GLR structure is similar to that of animal ionotropic glutamate receptors and is composed of four membrane-localized domains among which M2 is predicted not to span the membrane. Two glutamate binding domains (GlnH) are localized on the outside of the membrane.

Topology Models of Systems Catalyzing Ca2+ Efflux from the Cytosol. (A) Ca2+/H+ antiporters. The topology of CAX1 is based on speculation from hydropathy analyses. The number of transmembrane domains predicted varies from eight to eleven. CAX1 has recently been shown to have an N-terminal autoinhibitor. The blue highlight indicates the position of a 9-amino-acid sequence implicated in providing transport specificity for cations. (B) Ca2+-ATPase (ACA type). The topology of calcium pumps is well-supported by homology modeling based on a crystal structure of a mammalian sacro(endo)plasmic reticulum-type Ca2+ ATPase pump. ECA-type calcium pumps are predicted to have similar topologies, but lack the distinguishing feature of an N-terminal auotoinhibitor and calmodulin binding site.
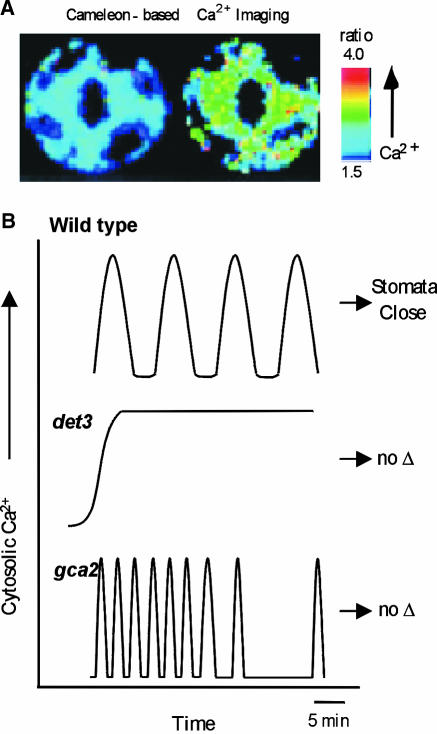
The Use of Arabidopsis Mutants and New Imaging Techniques Leads to a Better Understanding of Calcium Signaling. (A) Cameleon indicators expressed in guard cells allow a ratiometric measurement of cytosolic Ca2+ changes (low at left and high at right). (B) Mutants display abnormal Ca2+ signals that fail to trigger stomatal closure (Allen et al., 2000, 2001). In wild type (top), a variety of hormone or stress treatments trigger a series of Ca2+ oscillations that are followed by a prolonged closure of the stomata. In contrast, a steady state rise in calcium was observed in det3 mutants (middle) in response to treatments such as high external Ca2+ or oxidative stress (but not ABA and cold). Treatments that failed to produce normal oscillations also failed to trigger a prolonged stomatal closure. The stomatal closure response was also disrupted in the mutant gca2 (bottom). In this case, calcium signals occurred as an abnormal “high frequency” pattern of calcium oscillations. These results provide genetic evidence to support the hypothesis that a specific frequency of Ca2+ oscillations is required for a normal stomatal closure response.
References
-
- Allen, G.J., Chu, S.P., Harrington, C.L., Schumacher, K., Hoffman, T., Tang, Y.Y., Grill, E., and Schroeder, J.I. (2001). A defined range of guard cell calcium oscillation parameters encodes stomatal movements. Nature 411 1053–1057. - PubMed
-
- Allen, G.J., Chu, S.P., Schumacher, K., Shimazaki, C.T., Vafeados, D., Kemper, A., Hawke, S.D., Tallman, G., Tsien, R.Y., Harper, J.F., Chory, J., and Schroeder, J.I. (2000). Alteration of stimulus-specific guard cell calcium oscillations and stomatal closing in Arabidopsis det3 mutant. Science 289 2338–2342. - PubMed
-
- Allen, G.J., Kwak, J.M., Chu, S.P., Llopis, J., Tsien, R.Y., Harper, J.F., and Schroeder, J.I. (1999). Cameleon calcium indicator reports cytoplasmic calcium dynamics in Arabidopsis guard cells. Plant J. 19 735–747. - PubMed
-
- Allwood, E.G., Smertenko, A.P., and Hussey, P.J. (2001). Phosphorylation of plant actin-depolymerising factor by calmodulin-like domain protein kinase. FEBS Lett. 499 97–100. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
