

Dendritic K+ channels contribute to spike-timing dependent long-term potentiation in hippocampal pyramidal neurons
- PMID: 12048251
- PMCID: PMC123073
- DOI: 10.1073/pnas.122210599
Dendritic K+ channels contribute to spike-timing dependent long-term potentiation in hippocampal pyramidal neurons
Abstract
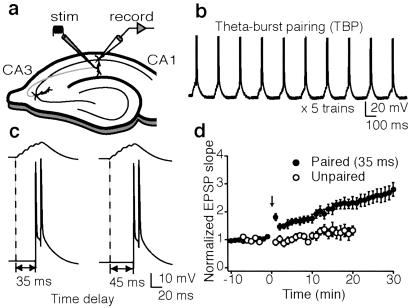
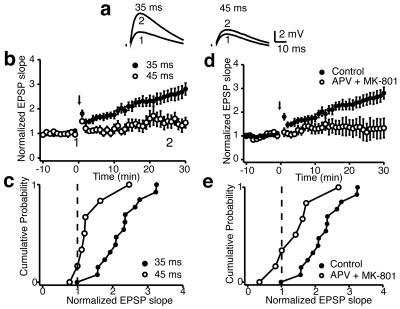
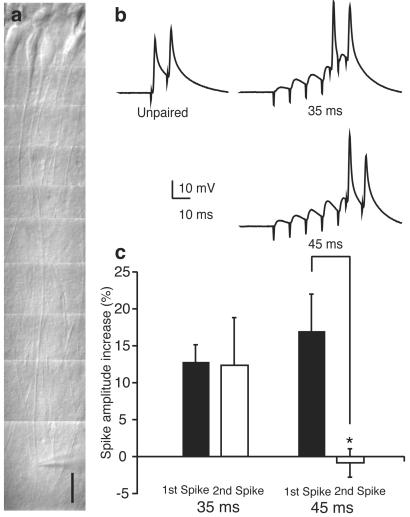
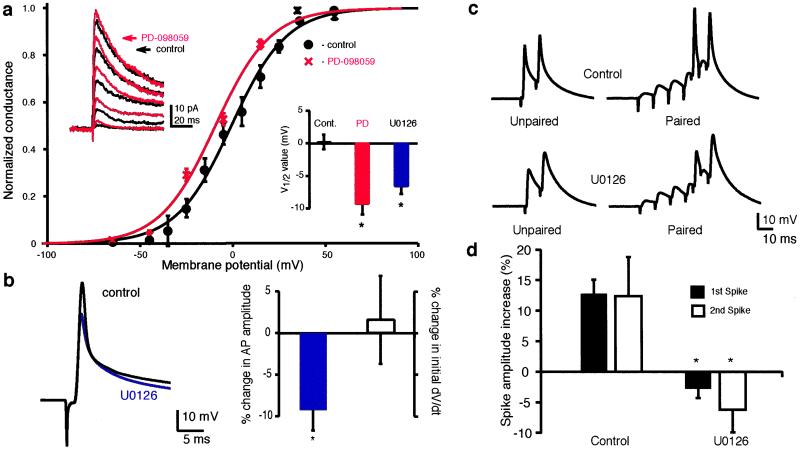
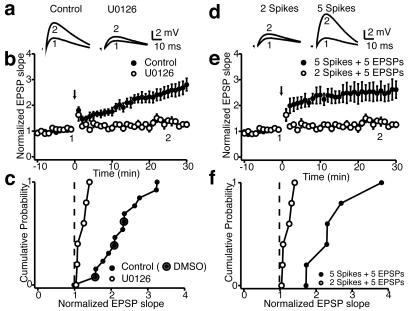
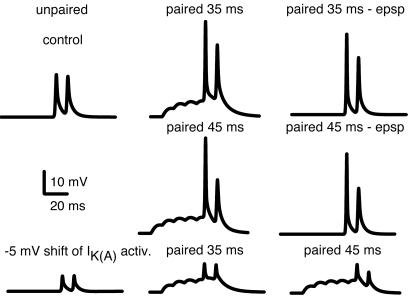
We investigated the role of A-type K(+) channels for the induction of long-term potentiation (LTP) of Schaffer collateral inputs to hippocampal CA1 pyramidal neurons. When low-amplitude excitatory postsynaptic potentials (EPSPs) were paired with two postsynaptic action potentials in a theta-burst pattern, N-methyl-d-aspartate (NMDA)-receptor-dependent LTP was induced. The amplitudes of the back-propagating action potentials were boosted in the dendrites only when they were coincident with the EPSPs. Mitogen-activated protein kinase (MAPK) inhibitors PD 098059 or U0126 shifted the activation of dendritic K(+) channels to more hyperpolarized potentials, reduced the boosting of dendritic action potentials by EPSPs, and suppressed the induction of LTP. These results support the hypothesis that dendritic K(+) channels and the boosting of back-propagating action potentials contribute to the induction of LTP in CA1 neurons.
Figures
Similar articles
-
Kinase-dependent modification of dendritic excitability after long-term potentiation.J Physiol. 2009 Jan 15;587(1):115-25. doi: 10.1113/jphysiol.2008.158816. Epub 2008 Nov 10. J Physiol. 2009. PMID: 19001050 Free PMC article.
-
Dendritic GIRK Channels Gate the Integration Window, Plateau Potentials, and Induction of Synaptic Plasticity in Dorsal But Not Ventral CA1 Neurons.J Neurosci. 2017 Apr 5;37(14):3940-3955. doi: 10.1523/JNEUROSCI.2784-16.2017. Epub 2017 Mar 9. J Neurosci. 2017. PMID: 28280255 Free PMC article.
-
A synaptically controlled, associative signal for Hebbian plasticity in hippocampal neurons.Science. 1997 Jan 10;275(5297):209-13. doi: 10.1126/science.275.5297.209. Science. 1997. PMID: 8985013
-
Active dendrites, potassium channels and synaptic plasticity.Philos Trans R Soc Lond B Biol Sci. 2003 Apr 29;358(1432):667-74. doi: 10.1098/rstb.2002.1248. Philos Trans R Soc Lond B Biol Sci. 2003. PMID: 12740112 Free PMC article. Review.
-
Dendritic potassium channels in hippocampal pyramidal neurons.J Physiol. 2000 May 15;525 Pt 1(Pt 1):75-81. doi: 10.1111/j.1469-7793.2000.00075.x. J Physiol. 2000. PMID: 10811726 Free PMC article. Review.
Cited by
-
Spatiotemporal characteristics of synaptic EPSP summation on the dendritic trees of hippocampal CA1 pyramidal neurons as revealed by laser uncaging stimulation.Cogn Neurodyn. 2011 Nov;5(4):333-42. doi: 10.1007/s11571-011-9158-9. Epub 2011 Jun 18. Cogn Neurodyn. 2011. PMID: 23115591 Free PMC article.
-
Neonatal Tissue Damage Promotes Spike Timing-Dependent Synaptic Long-Term Potentiation in Adult Spinal Projection Neurons.J Neurosci. 2016 May 11;36(19):5405-16. doi: 10.1523/JNEUROSCI.3547-15.2016. J Neurosci. 2016. PMID: 27170136 Free PMC article.
-
Tau-dependent Kv4.2 depletion and dendritic hyperexcitability in a mouse model of Alzheimer's disease.J Neurosci. 2015 Apr 15;35(15):6221-30. doi: 10.1523/JNEUROSCI.2552-14.2015. J Neurosci. 2015. PMID: 25878292 Free PMC article.
-
Signal propagation in oblique dendrites of CA1 pyramidal cells.J Neurophysiol. 2005 Dec;94(6):4145-55. doi: 10.1152/jn.00521.2005. J Neurophysiol. 2005. PMID: 16293591 Free PMC article.
-
Properties and functional role of voltage-dependent potassium channels in dendrites of rat cerebellar Purkinje neurons.J Neurosci. 2003 Jul 2;23(13):5698-707. doi: 10.1523/JNEUROSCI.23-13-05698.2003. J Neurosci. 2003. PMID: 12843273 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
