

Saccharomyces Rrm3p, a 5' to 3' DNA helicase that promotes replication fork progression through telomeric and subtelomeric DNA
- PMID: 12050116
- PMCID: PMC186315
- DOI: 10.1101/gad.982902
Saccharomyces Rrm3p, a 5' to 3' DNA helicase that promotes replication fork progression through telomeric and subtelomeric DNA
Abstract
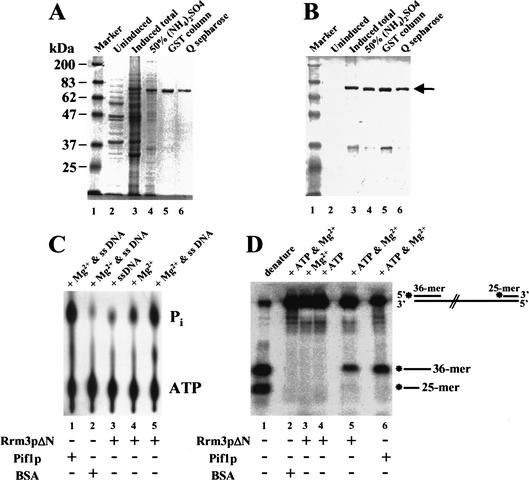
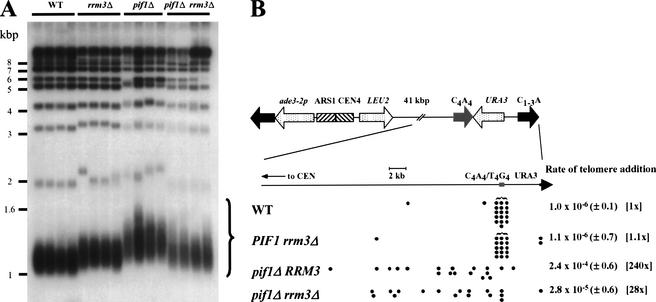
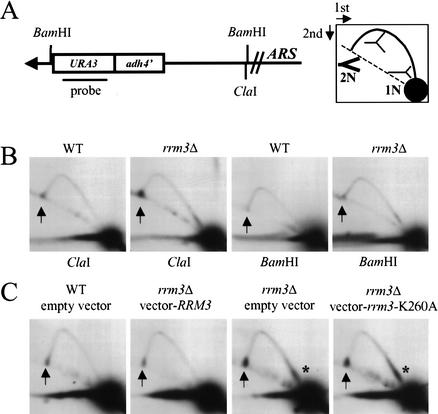
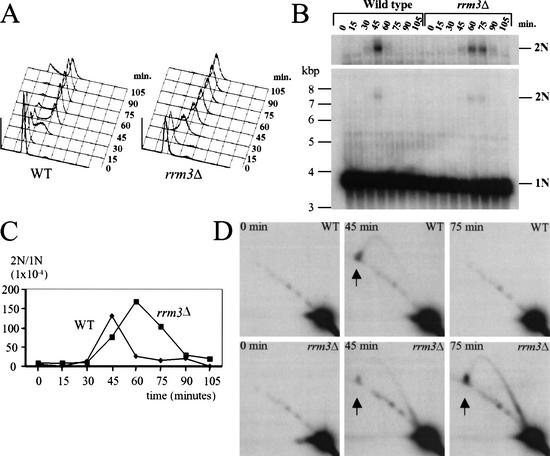
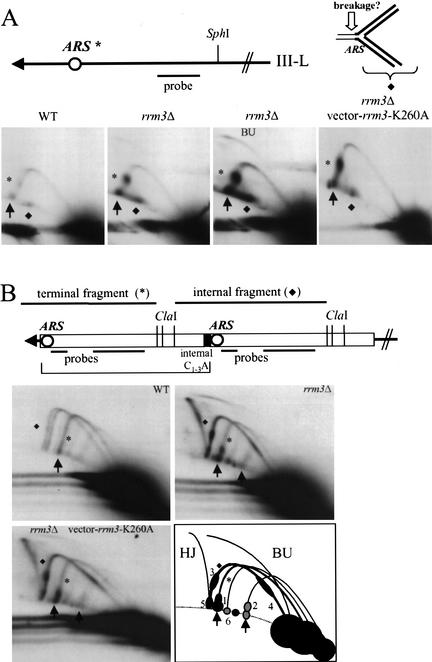
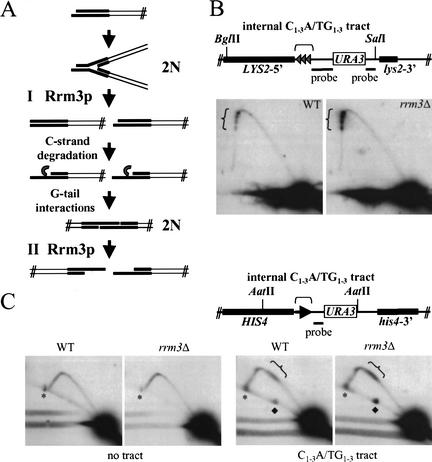
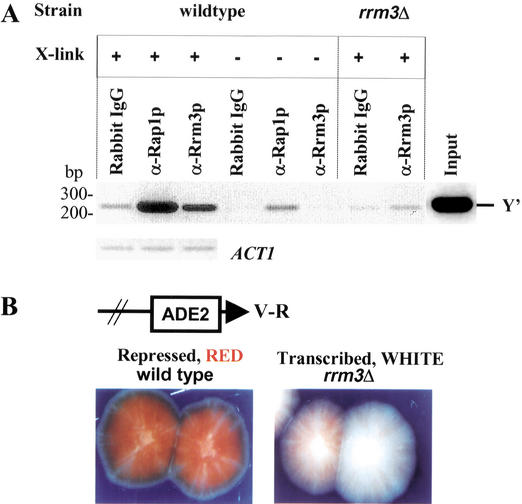
In wild-type Saccharomyces cerevisiae, replication forks slowed during their passage through telomeric C(1-3)A/TG(1-3) tracts. This slowing was greatly exacerbated in the absence of RRM3, shown here to encode a 5' to 3' DNA helicase. Rrm3p-dependent fork progression was seen at a modified Chromosome VII-L telomere, at the natural X-bearing Chromosome III-L telomere, and at Y'-bearing telomeres. Loss of Rrm3p also resulted in replication fork pausing at specific sites in subtelomeric DNA, such as at inactive replication origins, and at internal tracts of C(1-3)A/TG(1-3) DNA. The ATPase/helicase activity of Rrm3p was required for its role in telomeric and subtelomeric DNA replication. Because Rrm3p was telomere-associated in vivo, it likely has a direct role in telomere replication.
Figures
References
-
- Bennett RJ, Sharp JA, Wang JC. Purification and characterization of the Sgs1 DNA helicase activity of Saccharomyces cerevisiae. J Biol Chem. 1998;273:9644–9650. - PubMed
-
- Bessler JB, Torres JZ, Zakian VA. The Pif1p subfamily of helicases: Region specific DNA helicases. Trends Cell Biol. 2001;11:60–65. - PubMed
-
- Brewer BJ, Fangman WL. The localization of replication origins on ARS plasmids in S. cerevisiae. Cell. 1987;51:463–471. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous