

Involvement of both G(q/11) and G(s) proteins in gonadotropin-releasing hormone receptor-mediated signaling in L beta T2 cells
- PMID: 12050161
- PMCID: PMC2930616
- DOI: 10.1074/jbc.M203639200
Involvement of both G(q/11) and G(s) proteins in gonadotropin-releasing hormone receptor-mediated signaling in L beta T2 cells
Abstract
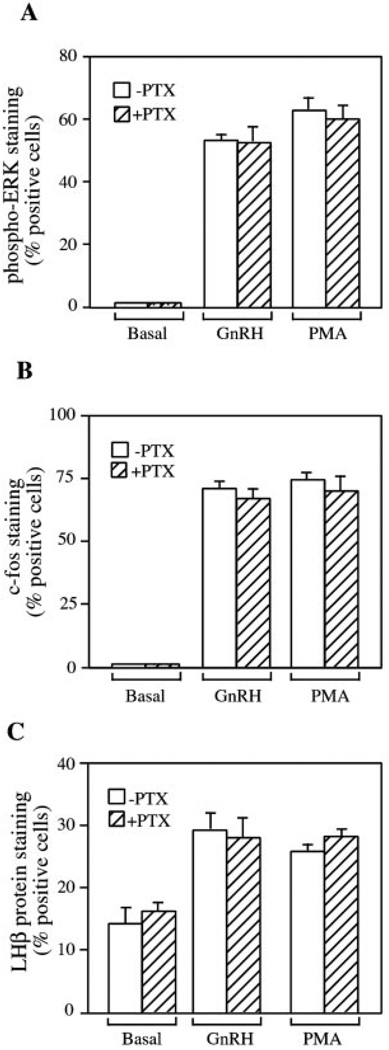
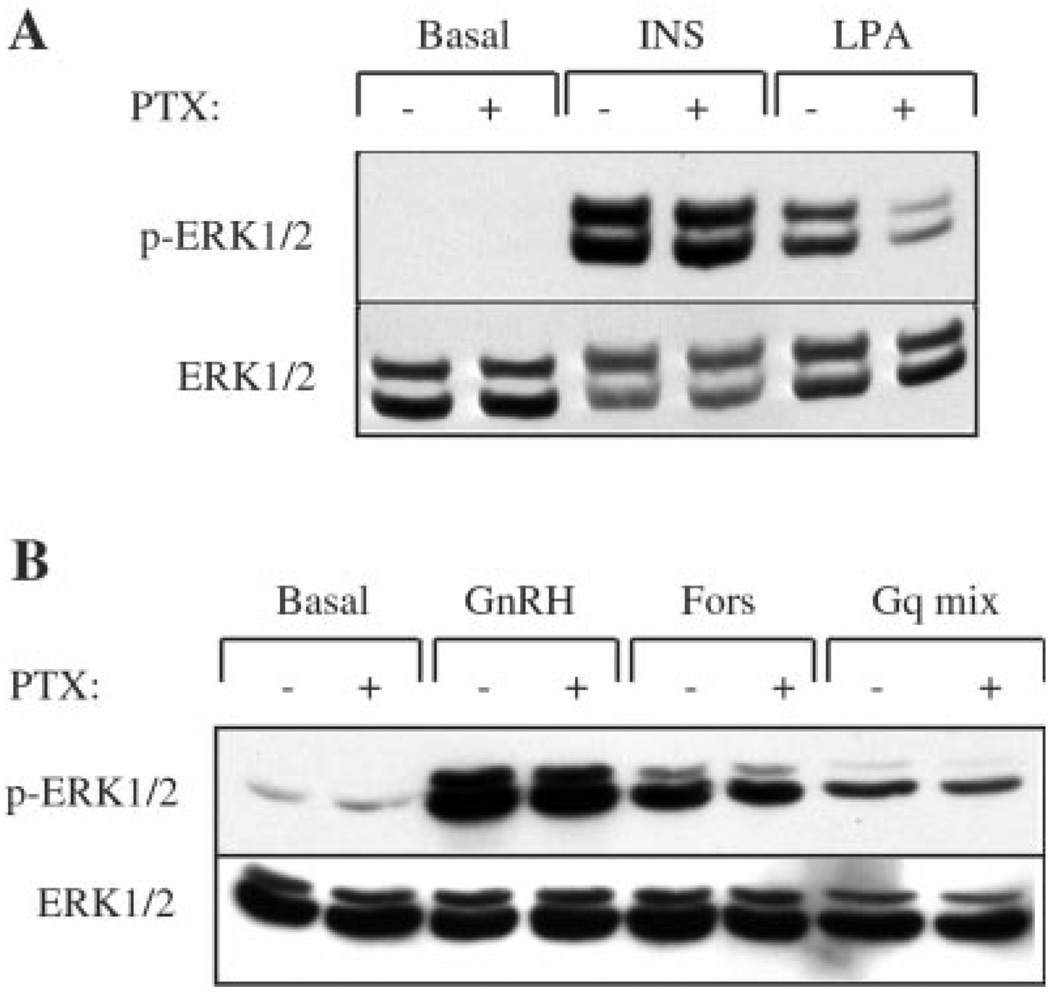
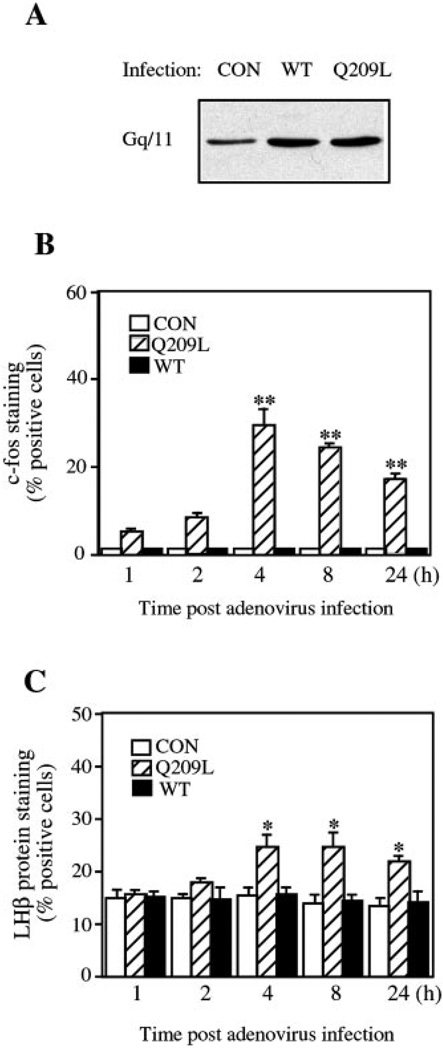
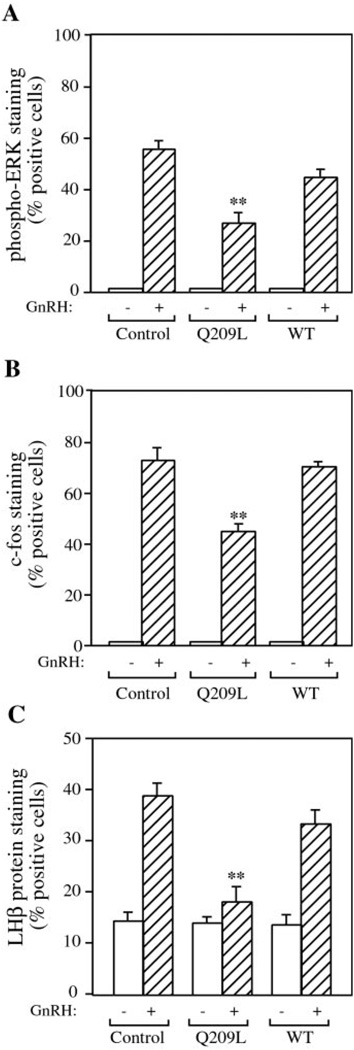
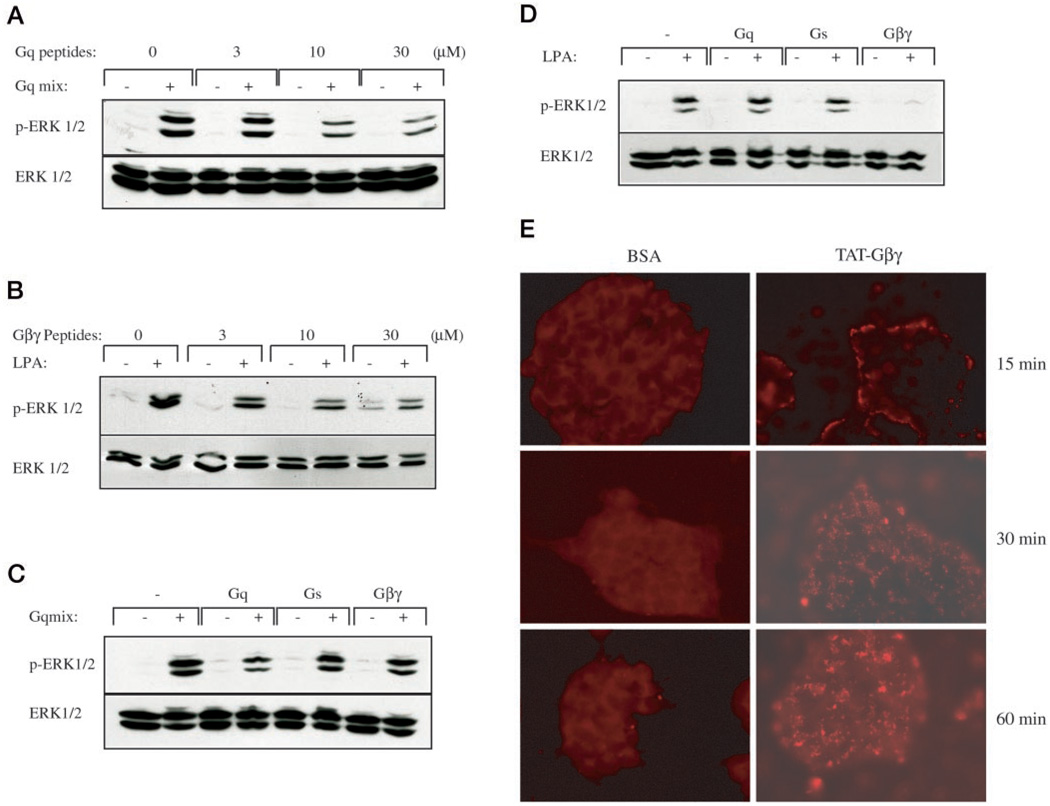
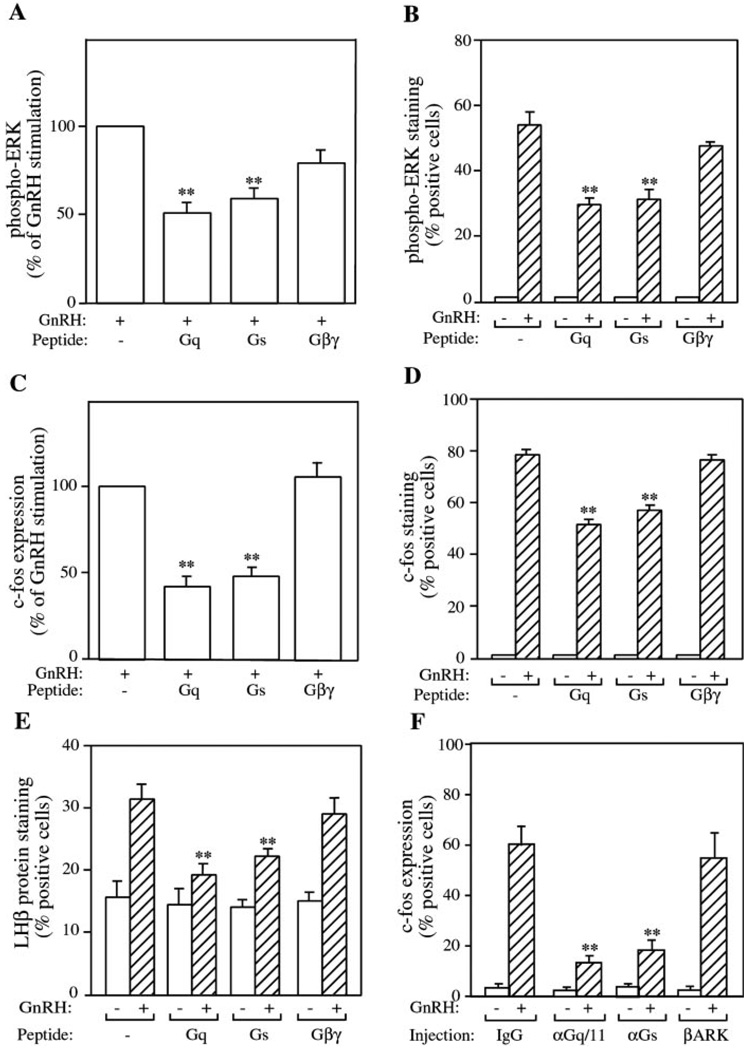
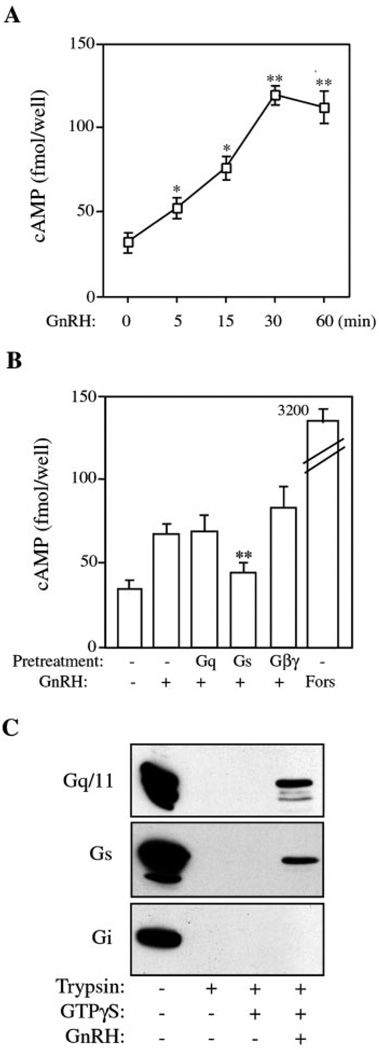
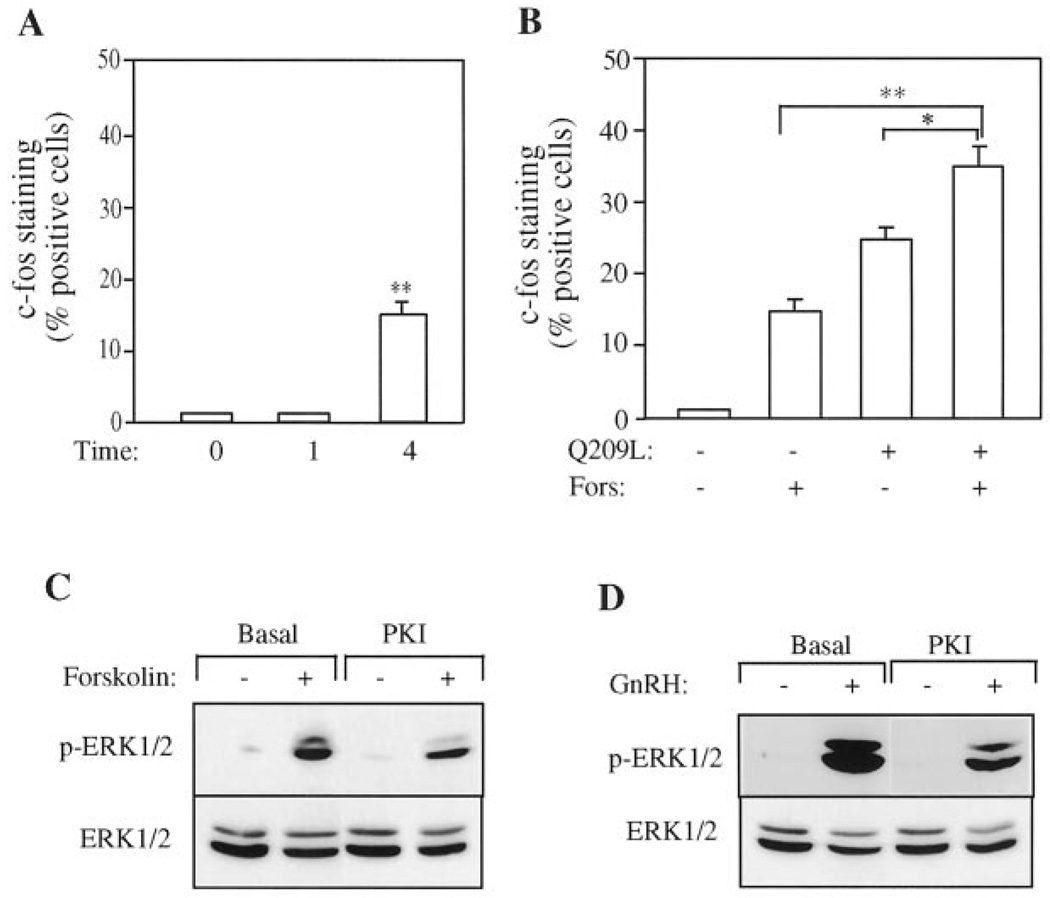
The hypothalamic hormone gonadotropin-releasing hormone (GnRH) stimulates the synthesis and release of the pituitary gonadotropins. GnRH acts through a plasma membrane receptor that is a member of the G protein-coupled receptor (GPCR) family. These receptors interact with heterotrimeric G proteins to initiate downstream signaling. In this study, we have investigated which G proteins are involved in GnRH receptor-mediated signaling in L beta T2 pituitary gonadotrope cells. We have shown previously that GnRH activates ERK and induces the c-fos and LH beta genes in these cells. Signaling via the G(i) subfamily of G proteins was excluded, as neither ERK activation nor c-Fos and LH beta induction was impaired by treatment with pertussis toxin or a cell-permeable peptide that sequesters G beta gamma-subunits. GnRH signaling was partially mimicked by adenoviral expression of a constitutively active mutant of G alpha(q) (Q209L) and was blocked by a cell-permeable peptide that uncouples G alpha(q) from GPCRs. Furthermore, chronic activation of G alpha(q) signaling induced a state of GnRH resistance. A cell-permeable peptide that uncouples G alpha(s) from receptors was also able to inhibit ERK, c-Fos, and LH beta, indicating that both G(q/11) and G(s) proteins are involved in signaling. Consistent with this, GnRH caused GTP loading on G(s) and G(q/11) and increased intracellular cAMP. Artificial elevation of cAMP with forskolin activated ERK and caused a partial induction of c-Fos. Finally, treatment of G alpha(q) (Q209L)-infected cells with forskolin enhanced the induction of c-Fos showing that the two pathways are independent and additive. Taken together, these results indicate that the GnRH receptor activates both G(q) and G(s) signaling to regulate gene expression in L beta T2 cells.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
