

Characterization of a spontaneous 9.5-kilobase-deletion mutant of murine gammaherpesvirus 68 reveals tissue-specific genetic requirements for latency
- PMID: 12050366
- PMCID: PMC136253
- DOI: 10.1128/jvi.76.13.6532-6544.2002
Characterization of a spontaneous 9.5-kilobase-deletion mutant of murine gammaherpesvirus 68 reveals tissue-specific genetic requirements for latency
Abstract
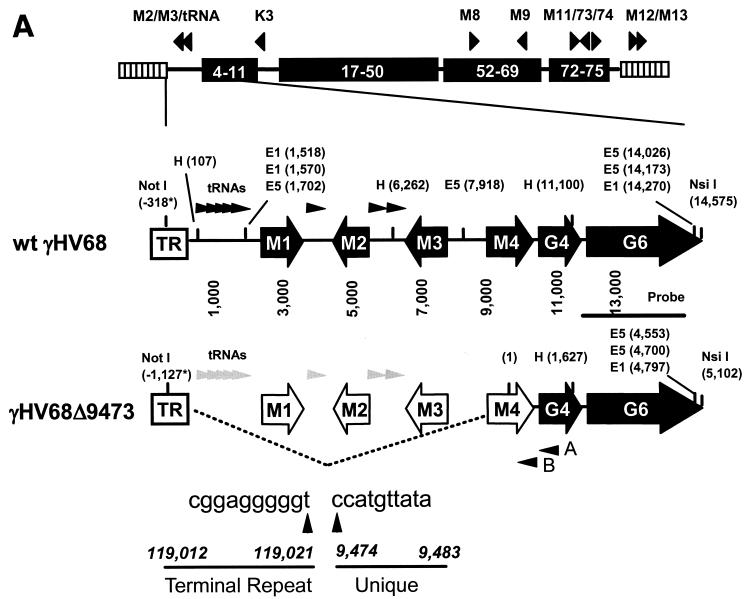
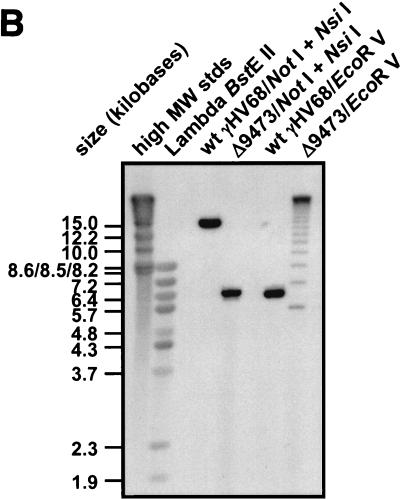
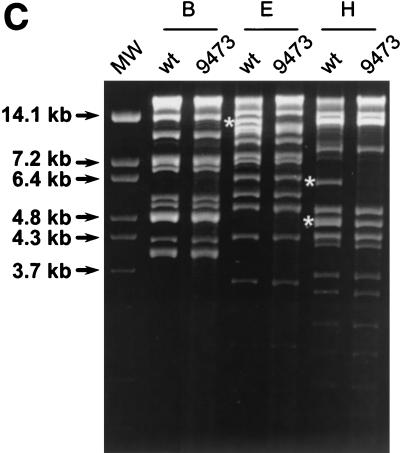
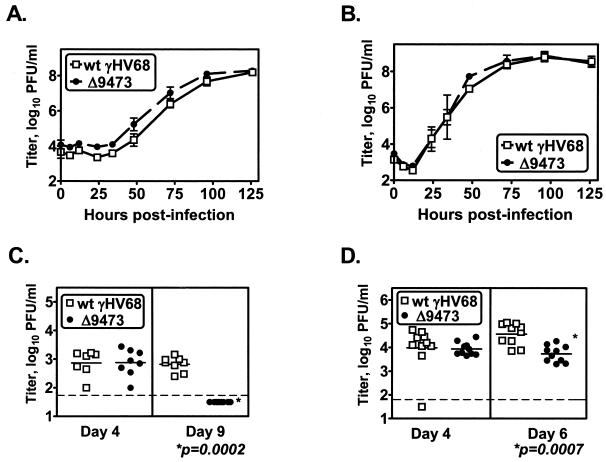
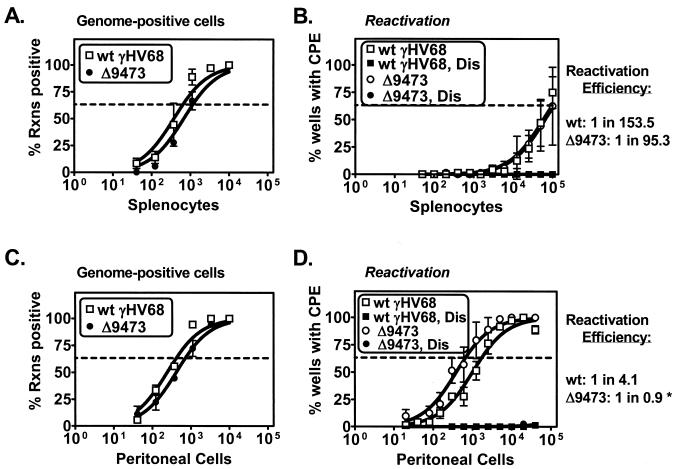
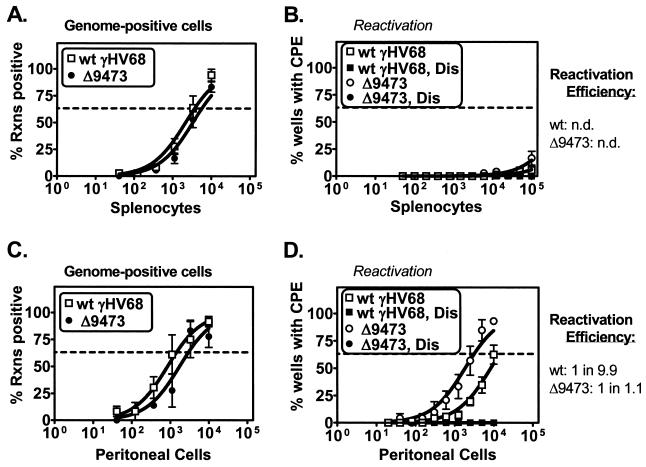
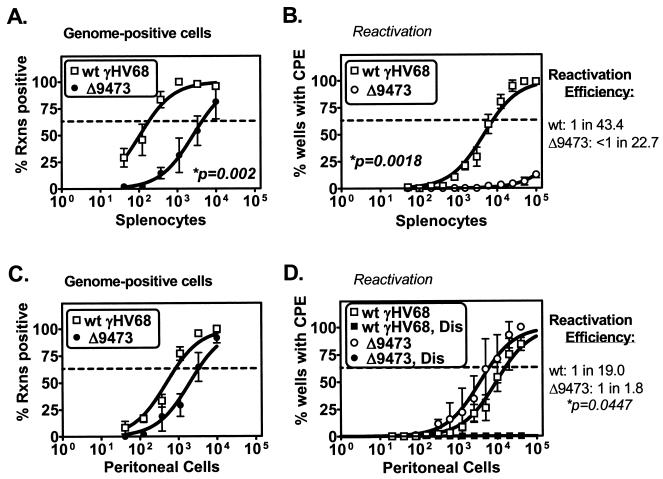
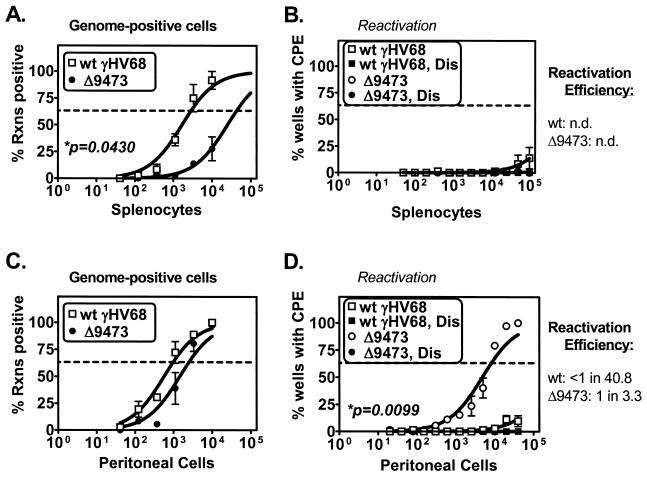
Murine gammaherpesvirus 68 (gammaHV68 [also known as MHV-68]) establishes a latent infection in mice, providing a small-animal model with which to identify host and viral factors that regulate gammaherpesvirus latency. While gammaHV68 establishes a latent infection in multiple tissues, including splenocytes and peritoneal cells, the requirements for latent infection within these tissues are poorly defined. Here we report the characterization of a spontaneous 9.5-kb-deletion mutant of gammaHV68 that lacks the M1, M2, M3, and M4 genes and eight viral tRNA-like genes. Previously, this locus has been shown to contain the latency-associated M2, M3, and viral tRNA-like genes. Through characterization of this mutant, we found that the M1, M2, M3, M4 genes and the viral tRNA-like genes are dispensable for (i) in vitro replication and (ii) the establishment and maintenance of latency in vivo and reactivation from latency following intraperitoneal infection. In contrast, following intranasal infection with this mutant, there was a defect in splenic latency at both early and late times, a phenotype not observed in peritoneal cells. These results indicate (i) that there are different genetic requirements for the establishment of latency in different latent reservoirs and (ii) that the genetic requirements for latency depend on the route of infection. While some of these phenotypes have been observed with specific mutations in the M1 and M2 genes, other phenotypes have never been observed with the available gammaHV68 mutants. These studies highlight the importance of loss-of-function mutations in defining the genetic requirements for the establishment and maintenance of herpesvirus latency.
Figures
References
-
- Babcock, G. J., L. L. Decker, M. Volk, and D. A. Thorley-Lawson. 1998. EBV persistence in memory B cells in vivo. Immunity 9:395-404. - PubMed
-
- Ballestas, M. E., P. A. Chatis, and K. M. Kaye. 1999. Efficient persistence of extrachromosomal KSHV DNA mediated by latency-associated nuclear antigen. Science 284:641-644. - PubMed
-
- Blaskovic, D., M. Stancekova, J. Svobodova, and J. Mistrikova. 1980. Isolation of five strains of herpesviruses from two species of free living small rodents. Acta Virol. 24:468. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
