

Critical role for glial cells in the propagation and spread of lymphocytic choriomeningitis virus in the developing rat brain
- PMID: 12050375
- PMCID: PMC136288
- DOI: 10.1128/jvi.76.13.6618-6635.2002
Critical role for glial cells in the propagation and spread of lymphocytic choriomeningitis virus in the developing rat brain
Abstract
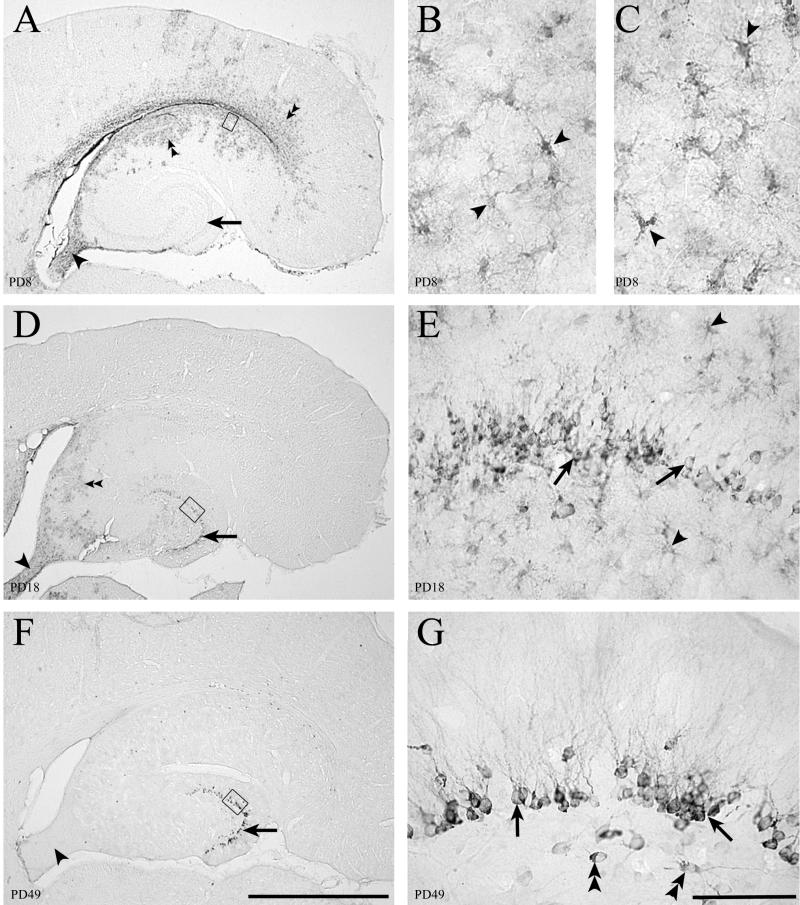
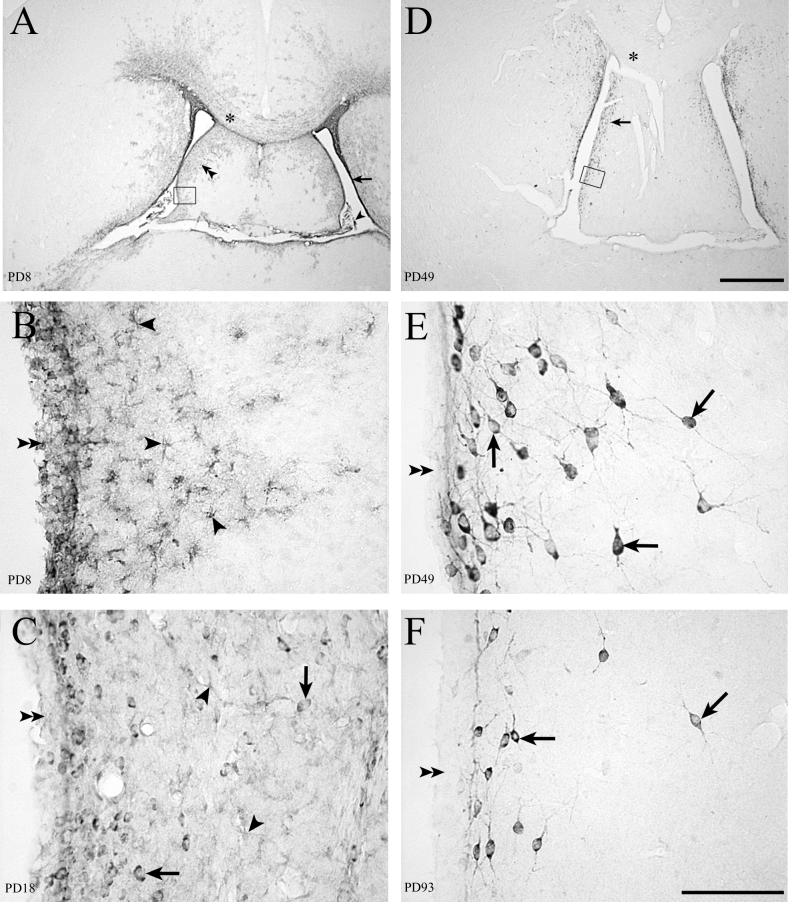
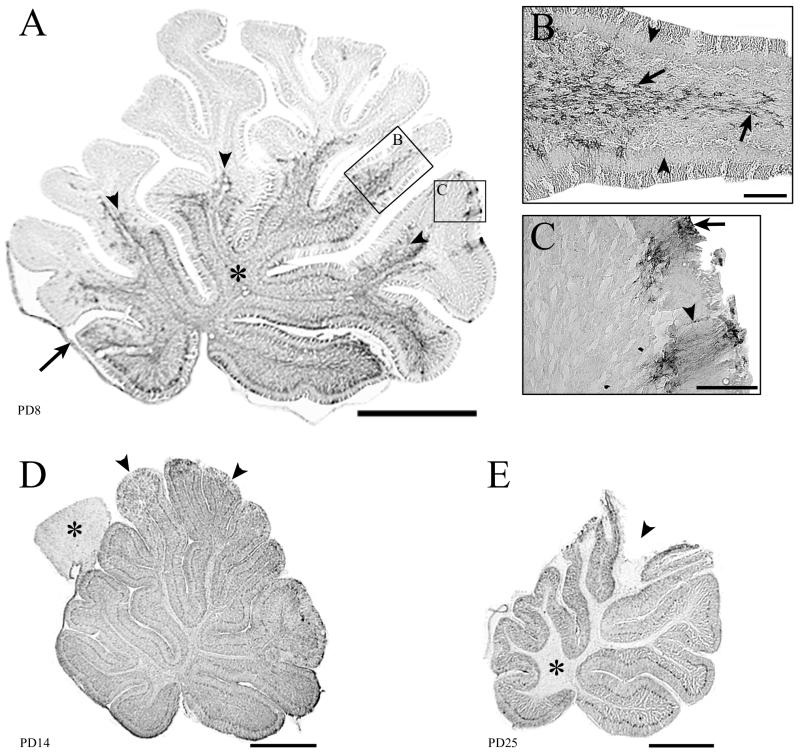
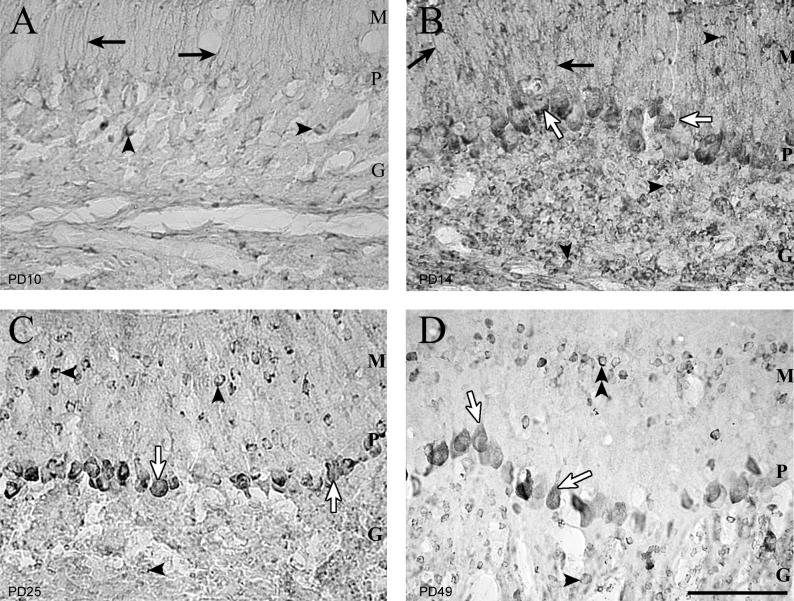
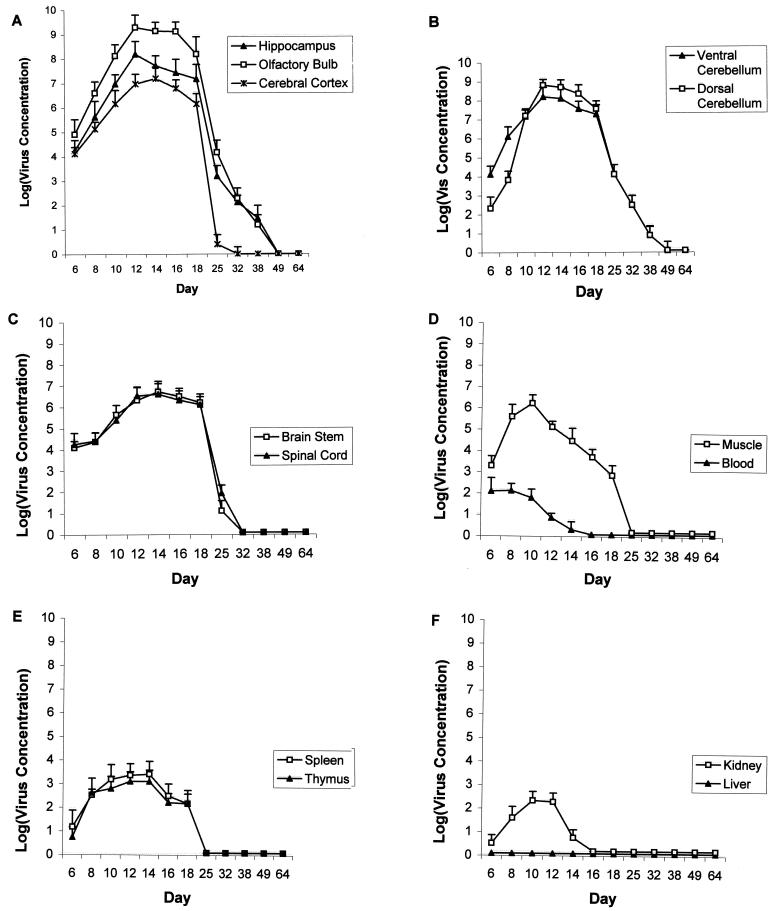
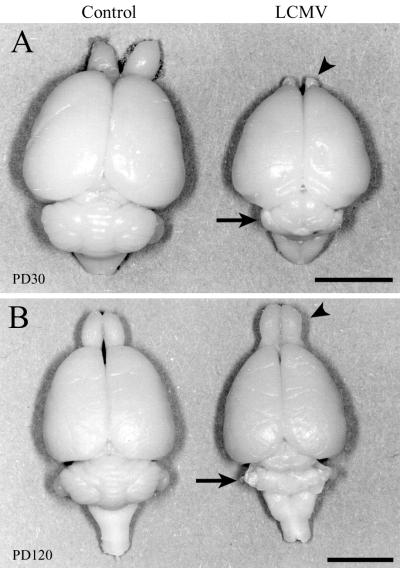
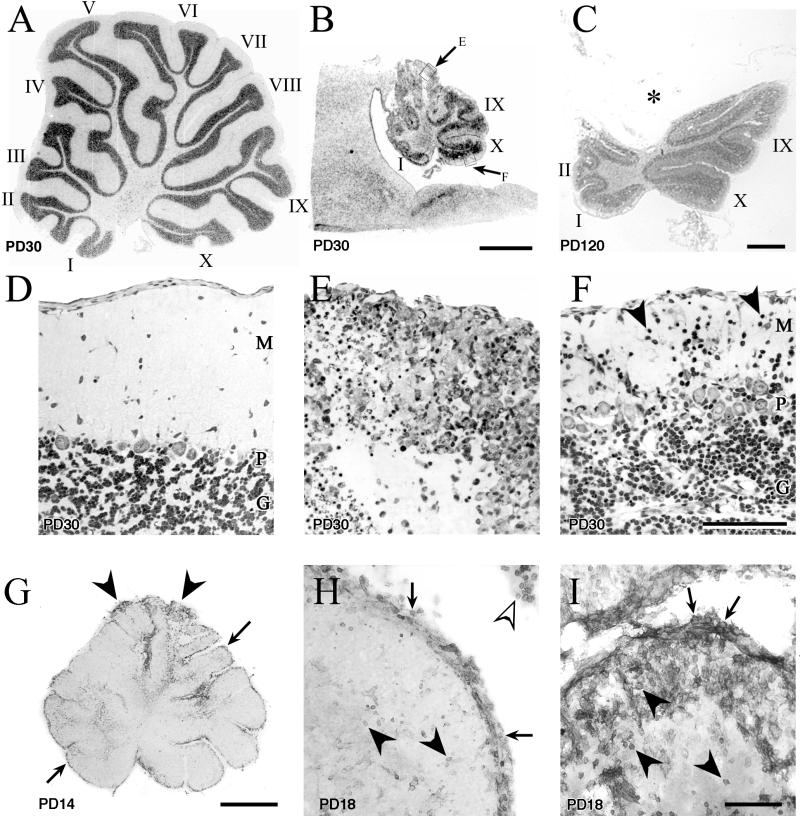
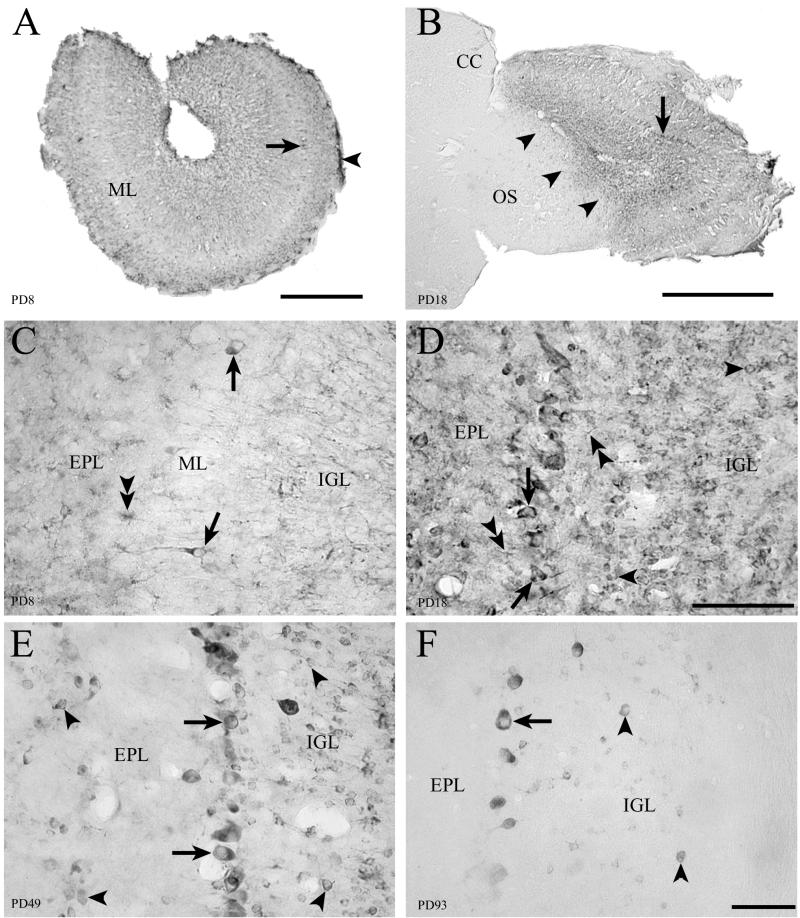
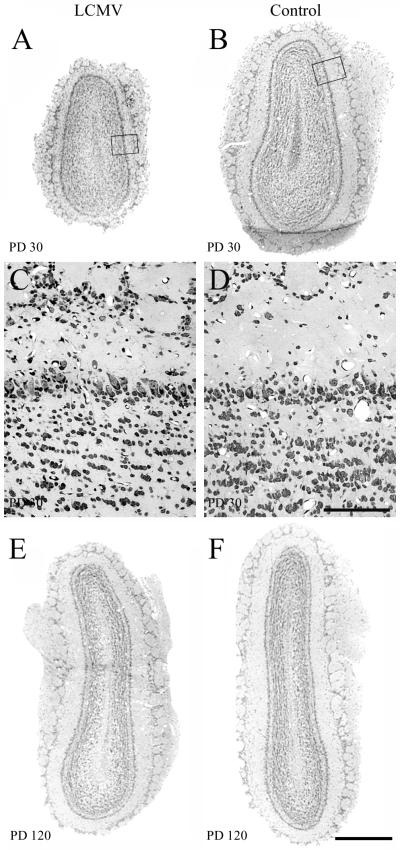
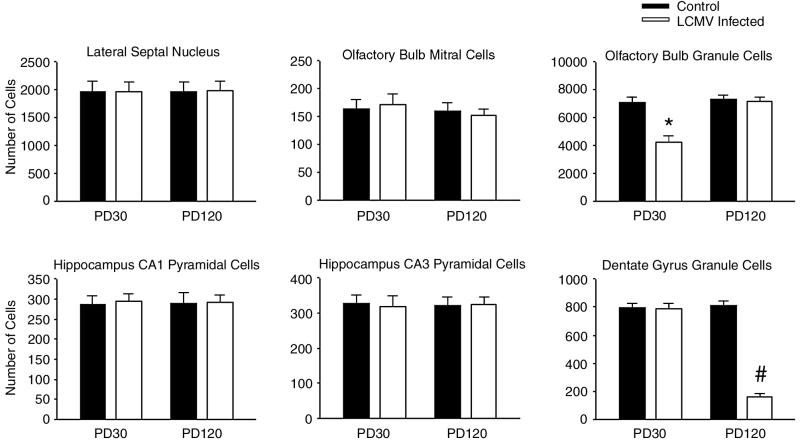
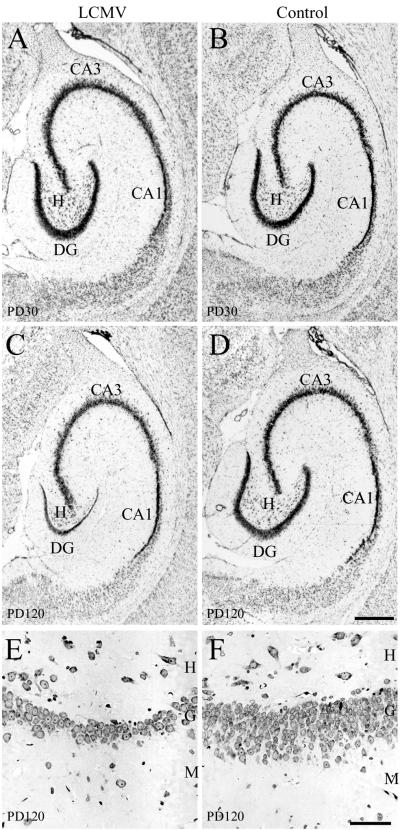
Inoculation of the neonatal rat with lymphocytic choriomeningitis virus (LCMV) results in the selective infection of several neuronal populations and in focal pathological changes. However, the pathway by which LCMV reaches the susceptible neurons has not been described, and the nature and time course of the pathological changes induced by the infection are largely unknown. This study examined the sequential migration of LCMV in the developing rat brain and compared the pathological changes among infected brain regions. The results demonstrate that astrocytes and Bergmann glia cells are the first cells of the brain parenchyma infected with LCMV and that the virus spreads across the brain principally via contiguous glial cells. The virus then spreads from glial cells into neurons. However, not all neurons are susceptible to infection. LCMV infects neurons in only four specific brain regions: the cerebellum, olfactory bulb, dentate gyrus, and periventricular region. The virus is then cleared from glial cells but persists in neurons. LCMV induces markedly different pathological changes in each of the four infected regions. The cerebellum undergoes an acute and permanent destruction, while the olfactory bulb is acutely hypoplastic but recovers fully with age. Neurons of the dentate gyrus are unaffected in the acute phase but undergo a delayed-onset mortality. In contrast, the periventricular region has neither acute nor late-onset cell loss. Thus, LCMV infects four specific brain regions in the developing brain by spreading from glial cells to neurons and then induces substantially different pathological changes with diverse time courses in each of the four infected regions.
Figures
References
-
- Altman, J. 1969. Autoradiographic and histological studies of postnatal neurogenesis. III. Dating the time of production and onset of differentiation of cerebellar microneurons in rats. J. Comp. Neurol. 136:269-294. - PubMed
-
- Altman, J. 1972. Postnatal development of the cerebellar cortex in the rat. II. Phases in the maturation of Purkinje cells and of the molecular layer. J. Comp. Neurol. 145:399-464. - PubMed
-
- Altman, J., and S. A. Bayer. 1997. Development of the cerebellar system, p. 498-523. CRC Press, New York, N.Y.
-
- Baldridge, J. R., B. D. Pearce, B. S. Parekh, and M. J. Buchmeier. 1993. Teratogenic effects of neonatal arenavirus infection on the developing rat cerebellum are abrogated by passive immunotherapy. J. Virol. 197:669-677. - PubMed
-
- Barton, L. L., S. C. Budd, W. S. Morfitt, C. J. Peters, T. G. Ksiazek, R. F. Schindler, and M. T. Yoshino. 1993. Congenital lymphocytic choriomeningitis virus infection in twins. Pediatr. Infect. Dis. J. 12:942-946. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
