

c-Src-mediated phosphorylation of hnRNP K drives translational activation of specifically silenced mRNAs
- PMID: 12052863
- PMCID: PMC133888
- DOI: 10.1128/MCB.22.13.4535-4543.2002
c-Src-mediated phosphorylation of hnRNP K drives translational activation of specifically silenced mRNAs
Abstract
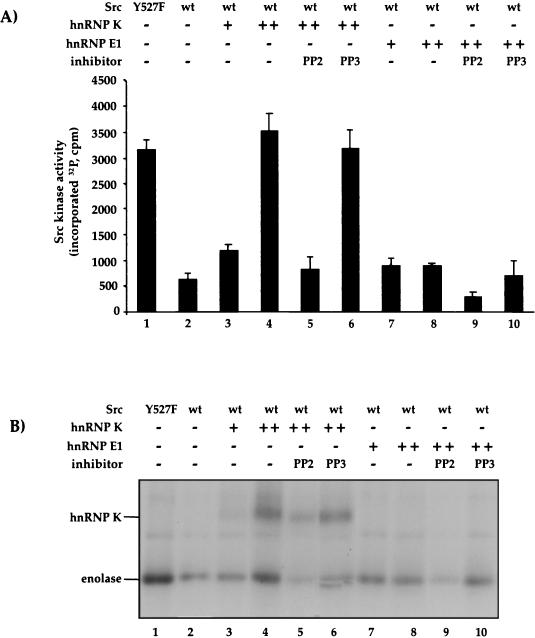
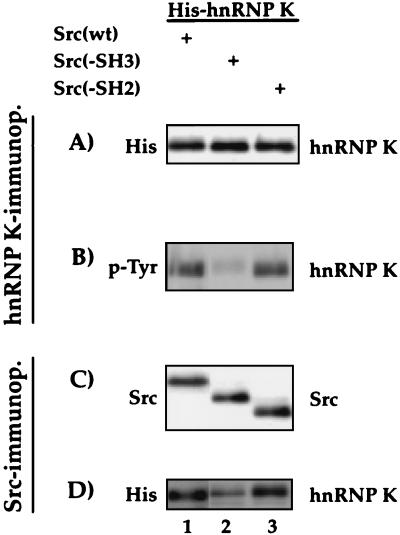
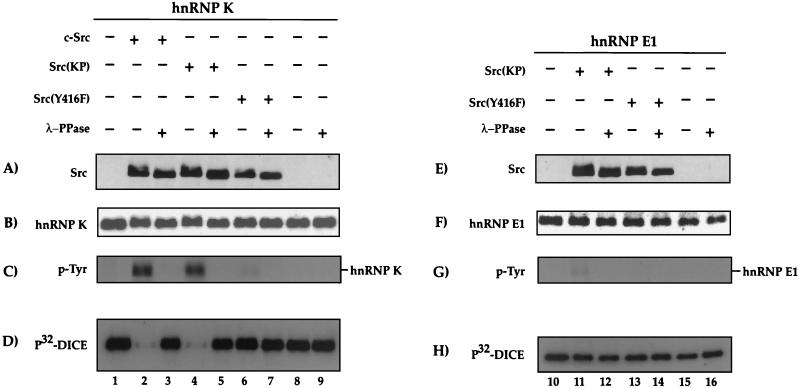
hnRNPK and hnRNP E1/E2 mediate translational silencing of cellular and viral mRNAs in a differentiation-dependent way by binding to specific regulatory sequences. The translation of 15-lipoxygenase (LOX) mRNA in erythroid precursor cells and of the L2 mRNA of human papilloma virus type 16 (HPV-16) in squamous epithelial cells is silenced when either of these cells is immature and is activated in maturing cells by unknown mechanisms. Here we address the question of how the silenced mRNA can be translationally activated. We show that hnRNP K and the c-Src kinase specifically interact with each other, leading to c-Src activation and tyrosine phosphorylation of hnRNP K in vivo and in vitro. c-Src-mediated phosphorylation reversibly inhibits the binding of hnRNP K to the differentiation control element (DICE) of the LOX mRNA 3' untranslated region in vitro and specifically derepresses the translation of DICE-bearing mRNAs in vivo. Our results establish a novel role of c-Src kinase in translational gene regulation and reveal a mechanism by which silenced mRNAs can be translationally activated.
Figures



References
-
- Alexandropoulos, K., and D. Baltimore. 1996. Coordinate activation of c-Src by SH3- and SH2-binding sites on a novel, p130 Cas-related protein, Sin. Genes Dev. 10:1341-1355. - PubMed
-
- Brasier, A. R., J. E. Tate, and J. F. Habener. 1989. Optimized use of the firefly luciferase assay as a reporter gene in mammalian cell lines. BioTechniques 7:1116-1122. - PubMed
-
- Brown, M. T., and J. A. Cooper. 1996. Regulation, substrates and functions of src. Biochim. Biophys. Acta 1287:121-149. - PubMed
-
- Collier, B., L. Goobar-Larsson, M. Sokolowski, and S. Schwartz. 1998. Translational inhibition in vitro of human papillomavirus type 16 L2 mRNA mediated through interaction with heterogenous ribonucleoprotein K and poly(rC)-binding protein 1 and 2. J. Biol. Chem. 273:22648-22656. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous