

ErbB2 is required for muscle spindle and myoblast cell survival
- PMID: 12052879
- PMCID: PMC133917
- DOI: 10.1128/MCB.22.13.4714-4722.2002
ErbB2 is required for muscle spindle and myoblast cell survival
Abstract
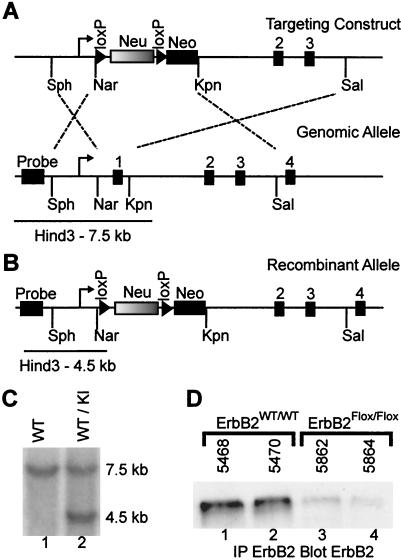
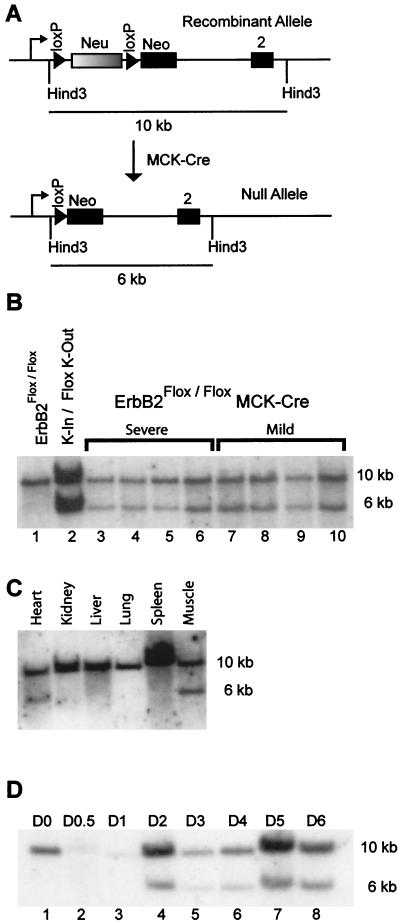
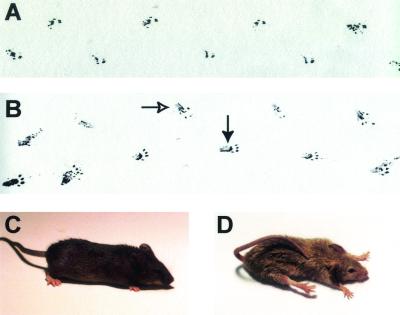
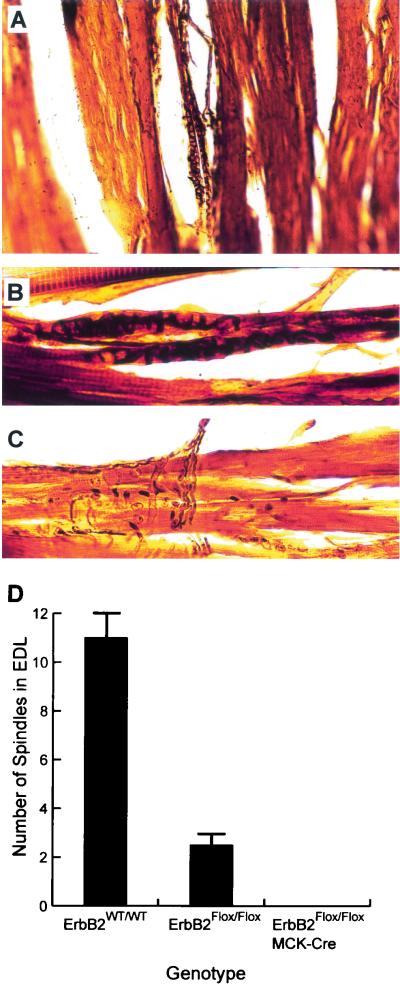
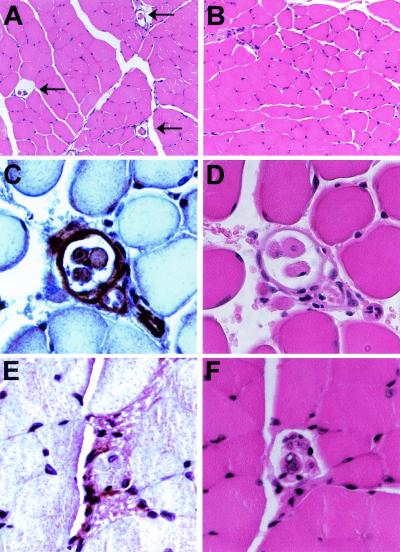
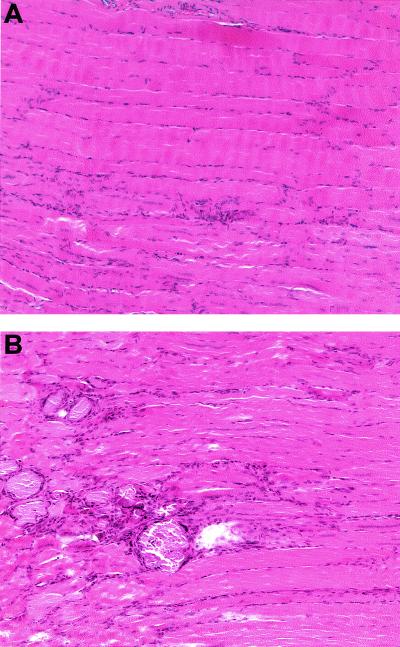
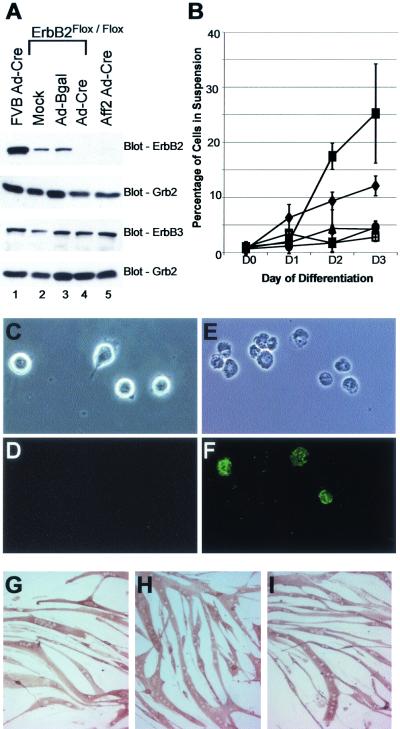
Signaling mediated by ErbB2 is thought to play a critical role in numerous developmental processes. However, due to the embryonic lethality associated with the germ line inactivation of erbB2, its role in adult tissues remains largely obscure. Given the expression of ErbB2 at the neuromuscular junction, we have created a muscle-specific knockout to assess its role there. This resulted in viable mice with a progressive defect in proprioception due to loss of muscle spindles. Interestingly, a partial reduction of ErbB2 levels also reduced the number of muscle spindles. Although histological analysis of the muscle revealed an otherwise normal architecture, induction of muscle injury revealed a defect in muscle regeneration. Consistent with these observations, primary myoblasts lacking ErbB2 exhibit extensive apoptosis upon differentiation into myofibers. Taken together, these results illustrate a dual role for ErbB2 in both muscle spindle maintenance and survival of myoblasts.
Figures
References
-
- Bargmann, C. I., M. C. Hung, and R. A. Weinberg. 1986. The neu oncogene encodes an epidermal growth factor receptor-related protein. Nature 319:226-230. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous