

Replication and subnuclear location dynamics of the immunoglobulin heavy-chain locus in B-lineage cells
- PMID: 12052893
- PMCID: PMC133899
- DOI: 10.1128/MCB.22.13.4876-4889.2002
Replication and subnuclear location dynamics of the immunoglobulin heavy-chain locus in B-lineage cells
Abstract
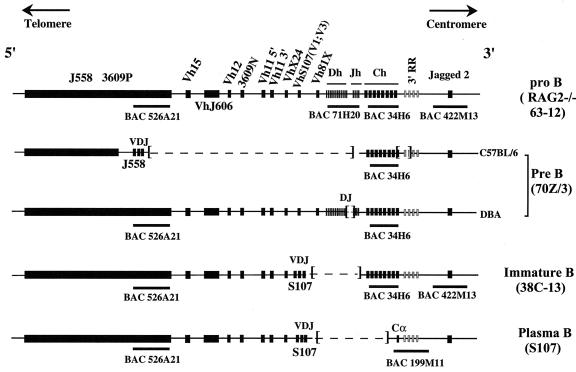
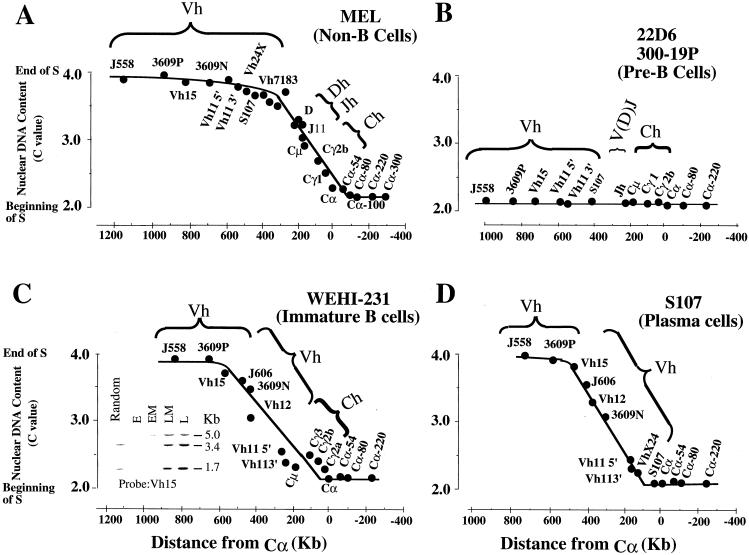
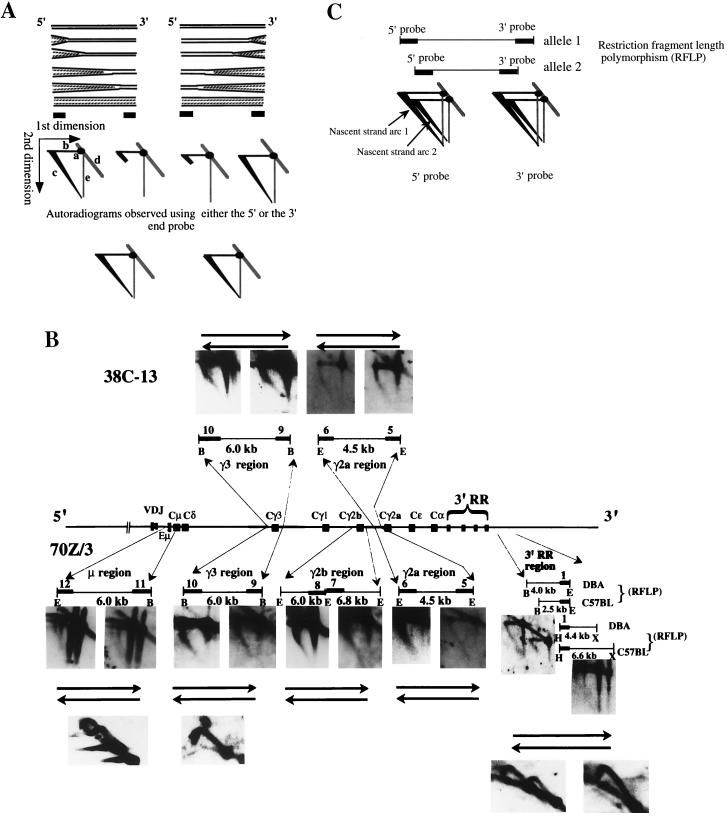
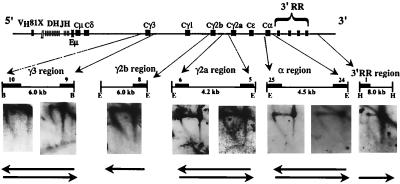
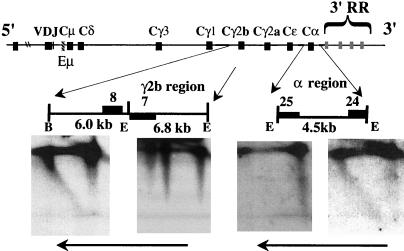
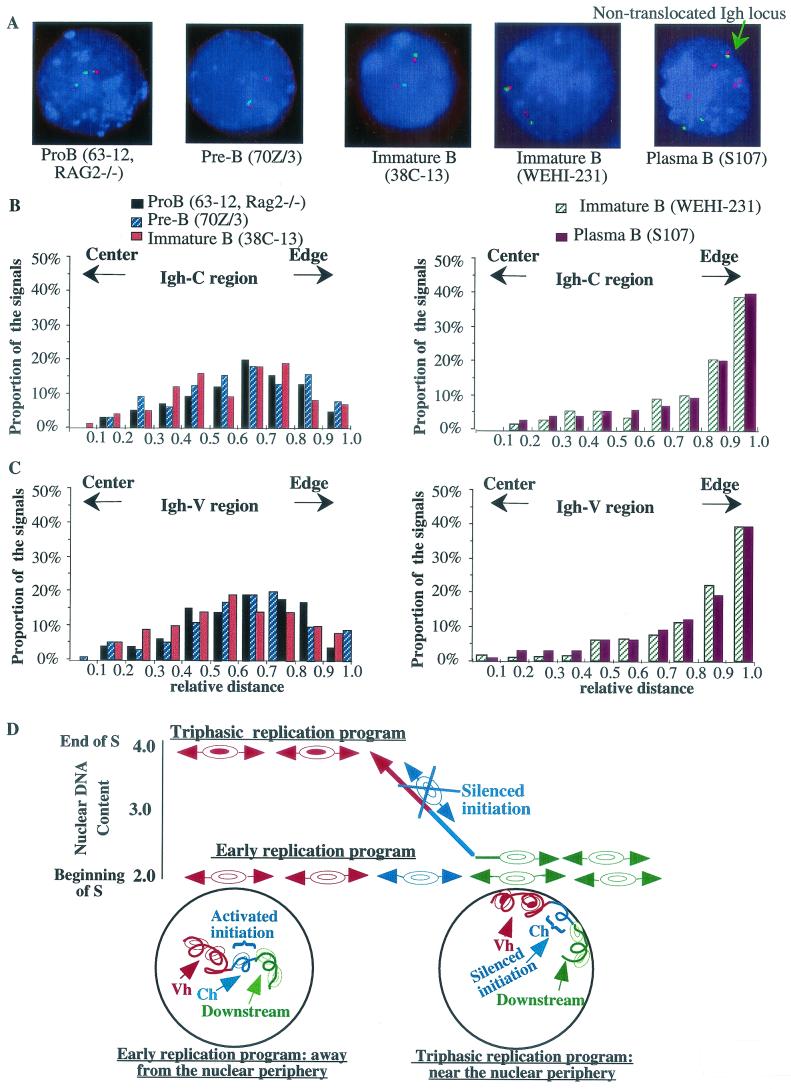
The murine immunoglobulin heavy-chain (Igh) locus provides an important model for understanding the replication of tissue-specific gene loci in mammalian cells. We have observed two DNA replication programs with dramatically different temporal replication patterns for the Igh locus in B-lineage cells. In pro- and pre-B-cell lines and in ex vivo-expanded pro-B cells, the entire locus is replicated early in S phase. In three cell lines that exhibit the early-replication pattern, we found that replication forks progress in both directions through the constant-region genes, which is consistent with the activation of multiple initiation sites. In contrast, in plasma cell lines, replication of the Igh locus occurs through a triphasic pattern similar to that previously detected in MEL cells. Sequences downstream of the Igh-C alpha gene replicate early in S, while heavy-chain variable (Vh) gene sequences replicate late in S. An approximately 500-kb transition region connecting sequences that replicate early and late is replicated progressively later in S. The formation of the transition region in different cell lines is independent of the sequences encompassed. In B-cell lines that exhibit a triphasic-replication pattern, replication forks progress in one direction through the examined constant-region genes. Timing data and the direction of replication fork movement indicate that replication of the transition region occurs by a single replication fork, as previously described for MEL cells. Associated with the contrasting replication programs are differences in the subnuclear locations of Igh loci. When the entire locus is replicated early in S, the Igh locus is located away from the nuclear periphery, but when Vh gene sequences replicate late and there is a temporal-transition region, the entire Igh locus is located near the nuclear periphery.
Figures
Similar articles
-
Changes in replication, nuclear location, and expression of the Igh locus after fusion of a pre-B cell line with a T cell line.J Immunol. 2005 Aug 15;175(4):2317-20. doi: 10.4049/jimmunol.175.4.2317. J Immunol. 2005. PMID: 16081801 Free PMC article.
-
Regulation of the replication of the murine immunoglobulin heavy chain gene locus: evaluation of the role of the 3' regulatory region.Mol Cell Biol. 1997 Oct;17(10):6167-74. doi: 10.1128/MCB.17.10.6167. Mol Cell Biol. 1997. PMID: 9315677 Free PMC article.
-
Evidence that a single replication fork proceeds from early to late replicating domains in the IgH locus in a non-B cell line.Mol Cell. 1999 Mar;3(3):321-30. doi: 10.1016/s1097-2765(00)80459-1. Mol Cell. 1999. PMID: 10198634
-
The IgH locus 3' regulatory region: pulling the strings from behind.Adv Immunol. 2011;110:27-70. doi: 10.1016/B978-0-12-387663-8.00002-8. Adv Immunol. 2011. PMID: 21762815 Review.
-
Temporal control of IgH gene expression in developing B cells by the 3' locus control region.Immunobiology. 1997 Dec;198(1-3):236-48. doi: 10.1016/S0171-2985(97)80044-7. Immunobiology. 1997. PMID: 9442395 Review.
Cited by
-
Changes in replication, nuclear location, and expression of the Igh locus after fusion of a pre-B cell line with a T cell line.J Immunol. 2005 Aug 15;175(4):2317-20. doi: 10.4049/jimmunol.175.4.2317. J Immunol. 2005. PMID: 16081801 Free PMC article.
-
DNA replication origins-where do we begin?Genes Dev. 2016 Aug 1;30(15):1683-97. doi: 10.1101/gad.285114.116. Genes Dev. 2016. PMID: 27542827 Free PMC article. Review.
-
Spatial distribution and specification of mammalian replication origins during G1 phase.J Cell Biol. 2003 Apr 28;161(2):257-66. doi: 10.1083/jcb.200211127. Epub 2003 Apr 21. J Cell Biol. 2003. PMID: 12707307 Free PMC article.
-
The temporal program of DNA replication: new insights into old questions.Chromosoma. 2006 Aug;115(4):273-87. doi: 10.1007/s00412-006-0062-8. Epub 2006 Mar 22. Chromosoma. 2006. PMID: 16552593 Review.
-
Developmentally regulated usage of Physarum DNA replication origins.EMBO Rep. 2003 May;4(5):474-8. doi: 10.1038/sj.embor.embor822. EMBO Rep. 2003. PMID: 12776736 Free PMC article.
References
-
- Alt, F., N. Rosenberg, S. Lewis, E. Thomas, and D. Baltimore. 1981. Organization and reorganization of immunoglobulin genes in A-MULV-transformed cells: rearrangement of heavy but not light chain genes. Cell 27:381-390. - PubMed
-
- Alwine, J. C. 1981. Hybridization analysis of specific RNAs by electrophoretic separation in agarose gels and transfer to diazobenzyloxymethyl paper. Gene Amplif. Anal. 2:565-572. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources