

Renal fibrosis. Extracellular matrix microenvironment regulates migratory behavior of activated tubular epithelial cells
- PMID: 12057905
- PMCID: PMC1850832
- DOI: 10.1016/S0002-9440(10)61150-9
Renal fibrosis. Extracellular matrix microenvironment regulates migratory behavior of activated tubular epithelial cells
Abstract
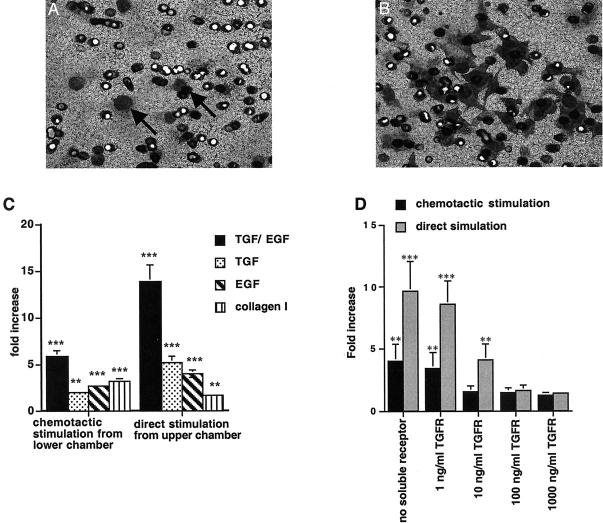
During progression of chronic renal disease, qualitative and quantitative changes in the composition of tubular basement membranes (TBMs) and interstitial matrix occur. Transforming growth factor (TGF)-beta(1)-mediated activation of tubular epithelial cells (TECs) is speculated to be a key contributor to the progression of tubulointerstitial fibrosis. To further understand the pathogenesis associated with renal fibrosis, we developed an in vitro Boyden chamber system using renal basement membranes that partially mimics in vivo conditions of TECs during health and disease. Direct stimulation of TECs with TGF-beta(1)/epithelial growth factor results in an increased migratory capacity across bovine TBM preparations. This is associated with increased matrix metalloproteinase (MMP) production, namely MMP-2 and MMP-9. Indirect chemotactic stimulation by TGF-beta(1)/EGF or collagen type I was insufficient in inducing migration of untreated TECs across bovine TBM preparation, suggesting that basement membrane integrity and composition play an important role in protecting TECs from interstitial fibrotic stimuli. Additionally, neutralization of MMPs by COL-3 inhibitor dramatically decreases the capacity of TGF-beta(1)-stimulated TECs to migrate through bovine TBM preparation. Collectively, these results demonstrate that basement membrane structure, integrity, and composition play an important role in determining interstitial influences on TECs and subsequent impact on potential aberrant cell-matrix interactions.
Figures




References
-
- Muller GA, Zeisberg M, Strutz F: The importance of tubulointerstitial damage in progressive renal disease. Nephrol Dial Transplant 2000, 15:76-77 - PubMed
-
- Remuzzi G, Bertani T: Pathophysiology of progressive nephropathies. N Engl J Med 1998, 339:1448-1456 - PubMed
-
- Abrass CK, Berfield AK, Stehman-Breen C, Alpers CE, Davis CL: Unique changes in interstitial extracellular matrix composition are associated with rejection and cyclosporine toxicity in human renal allograft biopsies. Am J Kidney Dis 1999, 33:11-20 - PubMed
-
- Brito PL, Fioretto P, Drummond K, Kim Y, Steffes MW, Basgen JM, Sisson-Ross S, Mauer M: Proximal tubular basement membrane width in insulin-dependent diabetes mellitus. Kidney Int 1998, 53:754-761 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
