

Roles for sigma factors in global circadian regulation of the cyanobacterial genome
- PMID: 12057947
- PMCID: PMC135120
- DOI: 10.1128/JB.184.13.3530-3538.2002
Roles for sigma factors in global circadian regulation of the cyanobacterial genome
Abstract
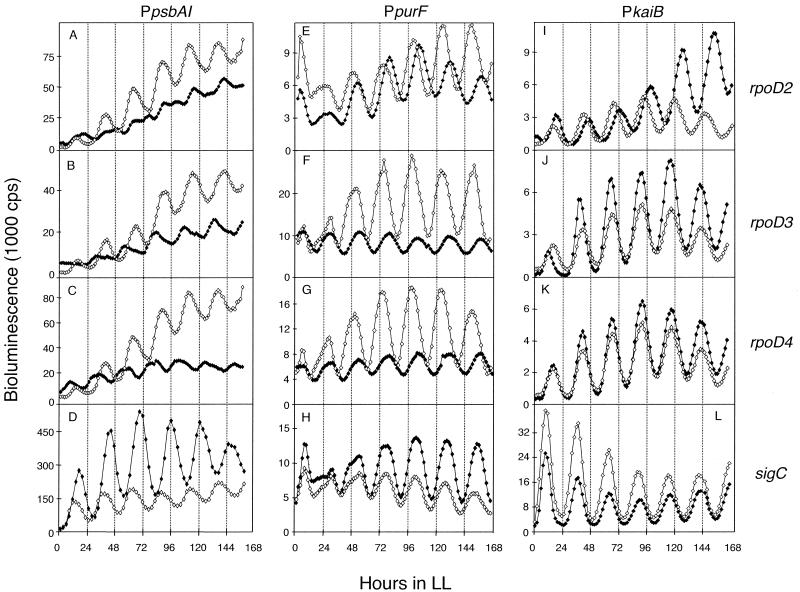
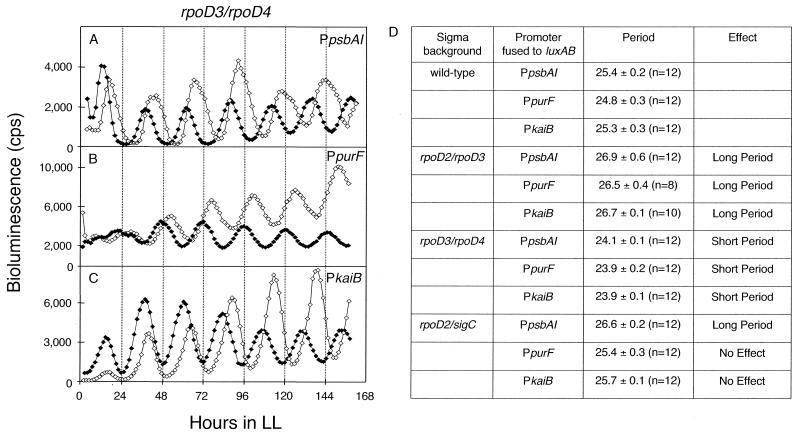
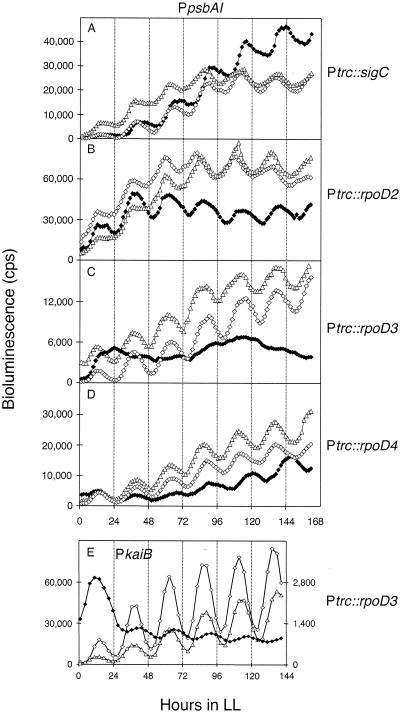
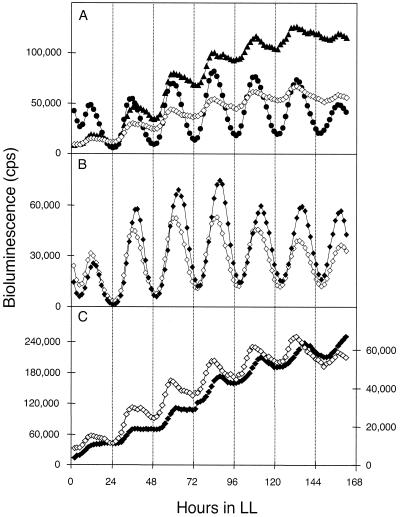
The circadian clock of the unicellular cyanobacterium Synechococcus elongatus PCC 7942 imposes a global rhythm of transcription on promoters throughout the genome. Inactivation of any of the four known group 2 sigma factor genes (rpoD2, rpoD3, rpoD4, and sigC), singly or pairwise, altered circadian expression from the psbAI promoter, changing amplitude, phase angle, waveform, or period. However, only the rpoD2 mutation and the rpoD3 rpoD4 and rpoD2 rpoD3 double mutations affected expression from the kaiB promoter. A striking differential effect was a 2-h lengthening of the circadian period of expression from the promoter of psbAI, but not of those of kaiB or purF, when sigC was inactivated. The data show that separate timing circuits with different periods can coexist in a cell. Overexpression of rpoD2, rpoD3, rpoD4, or sigC also changed the period or abolished the rhythmicity of PpsbAI expression, consistent with a model in which sigma factors work as a consortium to convey circadian information to downstream genes.
Figures
References
-
- Andersson, C. A., N. F. Tsinoremas, J. Shelton, N. V. Lebedeva, J. Yarrow, H. Min, and S. S. Golden. 2000. Application of bioluminescence to the study of circadian rhythms in cyanobacteria. Methods Enzymol. 305:527-542. - PubMed
-
- Bustos, S. A., and S. S. Golden. 1992. Light-regulated expression of the psbD gene family in Synechococcus sp. strain PCC 7942: evidence for the role of duplicated psbD genes in cyanobacteria. Mol. Gen. Genet. 232:221-230. - PubMed
-
- Clark, S. R., I. J. Oresnik, and M. F. Hynes. 2001. RpoN of Rhizobium leguminosarum bv. viciae strain VF39SM plays a central role in FnrN-dependent microaerobic regulation of genes involved in nitrogen fixation. Mol. Gen. Genet. 264:623-633. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
