

Kinesin-like protein CHO1 is required for the formation of midbody matrix and the completion of cytokinesis in mammalian cells
- PMID: 12058052
- PMCID: PMC117607
- DOI: 10.1091/mbc.01-10-0504
Kinesin-like protein CHO1 is required for the formation of midbody matrix and the completion of cytokinesis in mammalian cells
Abstract
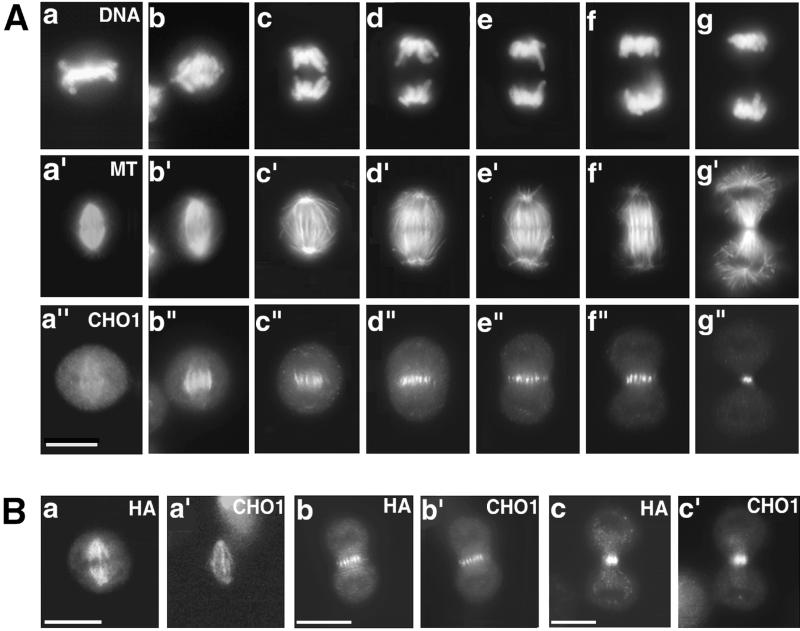
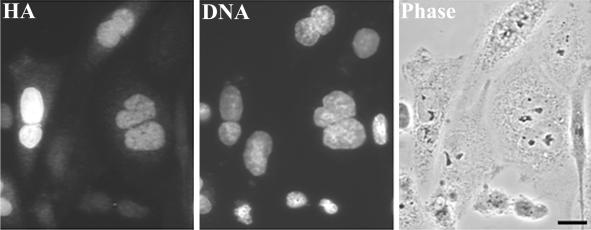
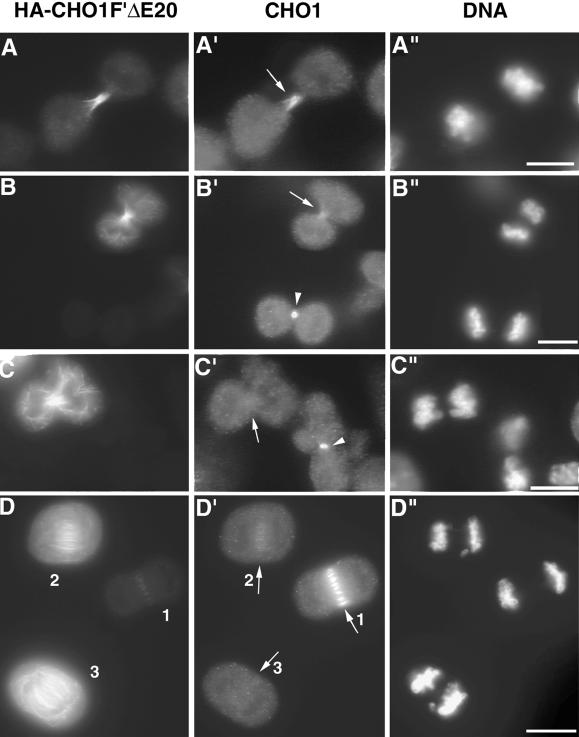
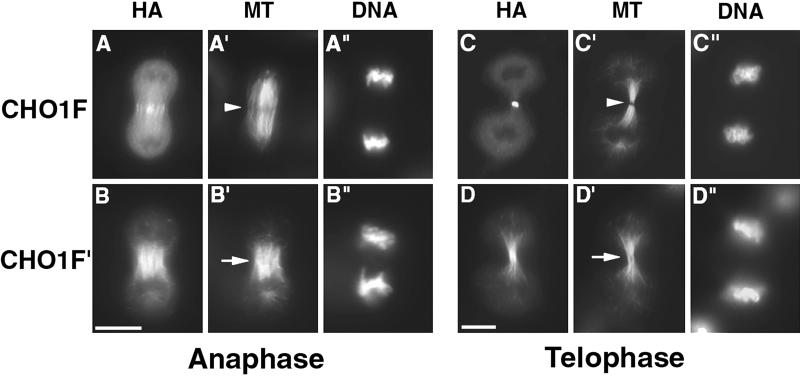
CHO1 is a mammalian kinesin-like motor protein of the MKLP1 subfamily. It associates with the spindle midzone during anaphase and concentrates to a midbody matrix during cytokinesis. CHO1 was originally implicated in karyokinesis, but the invertebrate homologues of CHO1 were shown to function in the midzone formation and cytokinesis. To analyze the role of the protein in mammalian cells, we mutated the ATP-binding site of CHO1 and expressed it in CHO cells. Mutant protein (CHO1F') was able to interact with microtubules via ATP-independent microtubule-binding site(s) but failed to accumulate at the midline of the central spindle and affected the localization of endogenous CHO1. Although the segregation of chromosomes, the bundling of midzone microtubules, and the initiation of cytokinesis proceeded normally in CHO1F'-expressing cells, the completion of cytokinesis was inhibited. Daughter cells were frequently entering interphase while connected by a microtubule-containing cytoplasmic bridge from which the dense midbody matrix was missing. Depletion of endogenous CHO1 via RNA-mediated interference also affected the formation of midbody matrix in dividing cells, caused the disorganization of midzone microtubules, and resulted in abortive cytokinesis. Thus, CHO1 may not be required for karyokinesis, but it is essential for the proper midzone/midbody formation and cytokinesis in mammalian cells.
Figures





Similar articles
-
Role of the midbody matrix in cytokinesis: RNAi and genetic rescue analysis of the mammalian motor protein CHO1.Mol Biol Cell. 2004 Jul;15(7):3083-94. doi: 10.1091/mbc.e03-12-0888. Epub 2004 Apr 9. Mol Biol Cell. 2004. PMID: 15075367 Free PMC article.
-
Recruitment of MKLP1 to the spindle midzone/midbody by INCENP is essential for midbody formation and completion of cytokinesis in human cells.Biochem J. 2005 Jul 15;389(Pt 2):373-81. doi: 10.1042/BJ20050097. Biochem J. 2005. PMID: 15796717 Free PMC article.
-
Molecular interactions of Polo-like-kinase 1 with the mitotic kinesin-like protein CHO1/MKLP-1.J Cell Sci. 2004 Jul 1;117(Pt 15):3233-46. doi: 10.1242/jcs.01173. Epub 2004 Jun 15. J Cell Sci. 2004. PMID: 15199097
-
Mechanisms of the Ase1/PRC1/MAP65 family in central spindle assembly.Biol Rev Camb Philos Soc. 2019 Dec;94(6):2033-2048. doi: 10.1111/brv.12547. Epub 2019 Jul 25. Biol Rev Camb Philos Soc. 2019. PMID: 31343816 Review.
-
Microtubules, membranes and cytokinesis.Curr Biol. 2000 Oct 19;10(20):R760-70. doi: 10.1016/s0960-9822(00)00746-6. Curr Biol. 2000. PMID: 11069103 Review.
Cited by
-
An ECT2-centralspindlin complex regulates the localization and function of RhoA.J Cell Biol. 2005 Aug 15;170(4):571-82. doi: 10.1083/jcb.200501097. J Cell Biol. 2005. PMID: 16103226 Free PMC article.
-
Kinesin 6 Regulation in Drosophila Female Meiosis by the Non-conserved N- and C- Terminal Domains.G3 (Bethesda). 2018 May 4;8(5):1555-1569. doi: 10.1534/g3.117.300571. G3 (Bethesda). 2018. PMID: 29514846 Free PMC article.
-
CYK4 relaxes the bias in the off-axis motion by MKLP1 kinesin-6.Commun Biol. 2021 Feb 10;4(1):180. doi: 10.1038/s42003-021-01704-2. Commun Biol. 2021. PMID: 33568771 Free PMC article.
-
Structural basis for Arf6-MKLP1 complex formation on the Flemming body responsible for cytokinesis.EMBO J. 2012 May 30;31(11):2590-603. doi: 10.1038/emboj.2012.89. Epub 2012 Apr 20. EMBO J. 2012. PMID: 22522702 Free PMC article.
-
The kinesin-like protein Pavarotti functions noncanonically to regulate actin dynamics.J Cell Biol. 2020 Sep 7;219(9):e201912117. doi: 10.1083/jcb.201912117. J Cell Biol. 2020. PMID: 32673395 Free PMC article.
References
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current protocols in molecular biology. New York: John Wiley & Sons, Inc; 1994.
-
- Boman AL, Kuai J, Zhu X, Chen J, Kuriyama R, Kahn RA. Arf proteins bind to mitotic kinesin-like protein 1 (MKLP1) in a GTP-dependent fashion. Cell Motil Cytoskeleton. 1999;44:119–132. - PubMed
-
- Chen M, Detrich HW., III The kinesin superfamily of zebrafish embryos. Mol Biol Cell. 1996;7:396a.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
