

Control of germination and lipid mobilization by COMATOSE, the Arabidopsis homologue of human ALDP
- PMID: 12065405
- PMCID: PMC125387
- DOI: 10.1093/emboj/cdf300
Control of germination and lipid mobilization by COMATOSE, the Arabidopsis homologue of human ALDP
Abstract
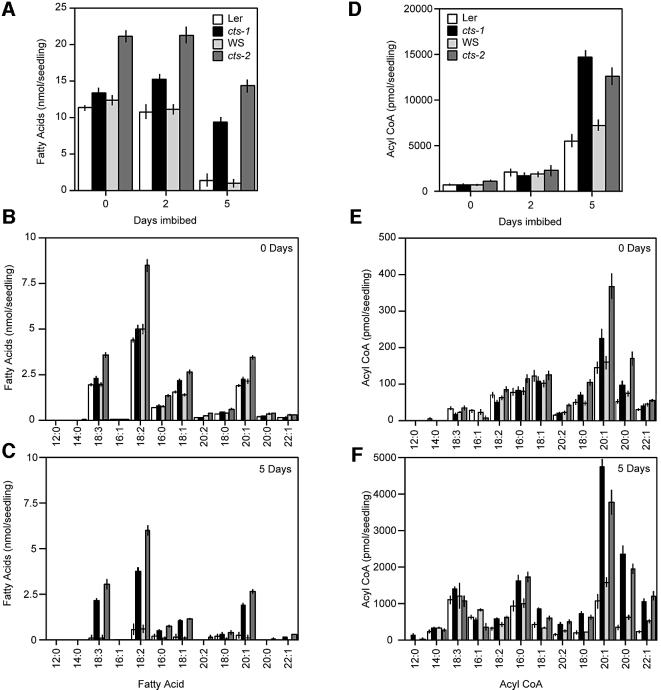
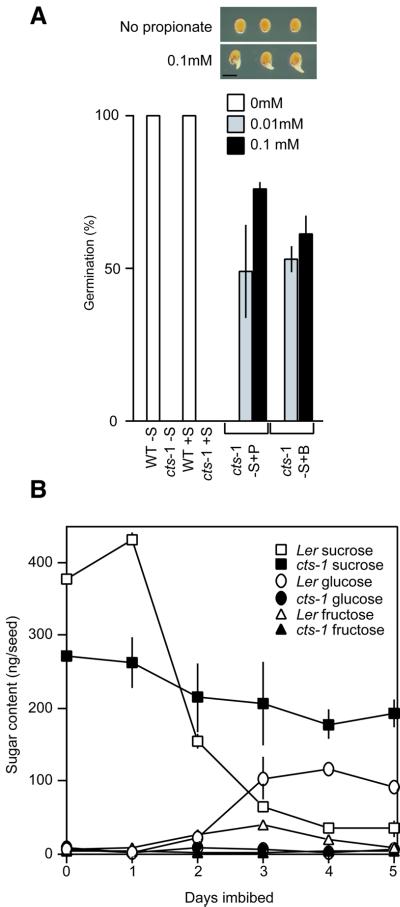
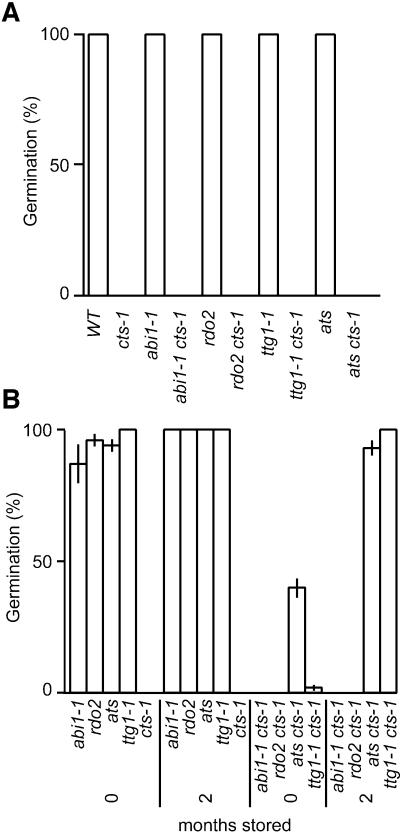
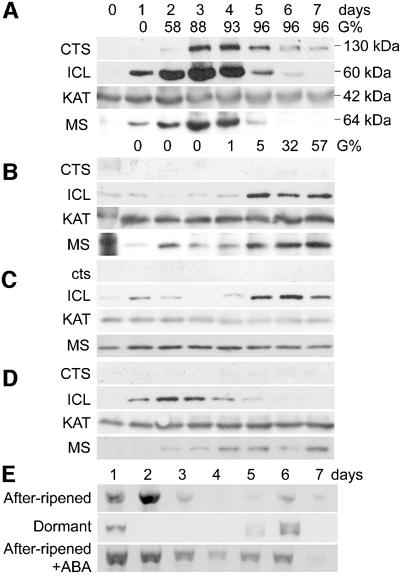
Embryo dormancy in flowering plants is an important dispersal mechanism that promotes survival of the seed through time. The subsequent transition to germination is a critical control point regulating initiation of vegetative growth. Here we show that the Arabidopsis COMATOSE (CTS) locus is required for this transition, and acts, at least in part, by profoundly affecting the metabolism of stored lipids. CTS encodes a peroxisomal protein of the ATP binding cassette (ABC) transporter class with significant identity to the human X-linked adrenoleukodystrophy protein (ALDP). Like X-ALD patients, cts mutant embryos and seedlings exhibit pleiotropic phenotypes associated with perturbation in fatty acid metabolism. CTS expression transiently increases shortly after imbibition during germination, but not in imbibed dormant seeds, and genetic analyses show that CTS is negatively regulated by loci that promote embryo dormancy through multiple independent pathways. Our results demonstrate that CTS regulates transport of acyl CoAs into the peroxisome, and indicate that regulation of CTS function is a major control point for the switch between the opposing developmental programmes of dormancy and germination.
Figures
Similar articles
-
Conservation of targeting but divergence in function and quality control of peroxisomal ABC transporters: an analysis using cross-kingdom expression.Biochem J. 2011 Jun 15;436(3):547-57. doi: 10.1042/BJ20110249. Biochem J. 2011. PMID: 21476988
-
Analysis of the role of COMATOSE and peroxisomal beta-oxidation in the determination of germination potential in Arabidopsis.J Exp Bot. 2006;57(11):2805-14. doi: 10.1093/jxb/erl045. Epub 2006 Jul 14. J Exp Bot. 2006. PMID: 16844736
-
The Arabidopsis ALDP protein homologue COMATOSE is instrumental in peroxisomal acetate metabolism.Biochem J. 2007 Sep 15;406(3):399-406. doi: 10.1042/BJ20070258. Biochem J. 2007. PMID: 17581114 Free PMC article.
-
X-linked adrenoleukodystrophy: very long-chain fatty acid metabolism, ABC half-transporters and the complicated route to treatment.Mol Genet Metab. 2007 Mar;90(3):268-76. doi: 10.1016/j.ymgme.2006.10.001. Epub 2006 Nov 7. Mol Genet Metab. 2007. PMID: 17092750 Review.
-
Peroxisomal ABC transporters: functions and mechanism.Biochem Soc Trans. 2015 Oct;43(5):959-65. doi: 10.1042/BST20150127. Epub 2015 Oct 9. Biochem Soc Trans. 2015. PMID: 26517910 Free PMC article. Review.
Cited by
-
Transport proteins regulate the flux of metabolites and cofactors across the membrane of plant peroxisomes.Front Plant Sci. 2012 Jan 16;3:3. doi: 10.3389/fpls.2012.00003. eCollection 2012. Front Plant Sci. 2012. PMID: 22645564 Free PMC article.
-
Acyl-lipid metabolism.Arabidopsis Book. 2010;8:e0133. doi: 10.1199/tab.0133. Epub 2010 Jun 11. Arabidopsis Book. 2010. PMID: 22303259 Free PMC article.
-
The Arabidopsis peroxisomal targeting signal type 2 receptor PEX7 is necessary for peroxisome function and dependent on PEX5.Mol Biol Cell. 2005 Feb;16(2):573-83. doi: 10.1091/mbc.e04-05-0422. Epub 2004 Nov 17. Mol Biol Cell. 2005. PMID: 15548601 Free PMC article.
-
Hydro-Electro Hybrid Priming Promotes Carrot (Daucus carota L.) Seed Germination by Activating Lipid Utilization and Respiratory Metabolism.Int J Mol Sci. 2021 Oct 14;22(20):11090. doi: 10.3390/ijms222011090. Int J Mol Sci. 2021. PMID: 34681749 Free PMC article.
-
Whole-genome survey of the putative ATP-binding cassette transporter family genes in Vitis vinifera.PLoS One. 2013 Nov 11;8(11):e78860. doi: 10.1371/journal.pone.0078860. eCollection 2013. PLoS One. 2013. PMID: 24244377 Free PMC article.
References
-
- Baskin C.C. and Baskin,J.M. (1998) Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. Academic Press, London, UK.
-
- Baumlein H., Misera,S., Luerssen,H., Kolle,K., Horstmann,C., Wobus,U. and Muller,A.J. (1994) The FUS3 gene of Arabidopsis thaliana is a regulator of gene expression during late embryogenesis. Plant J., 6, 379–387.
-
- Bewley J.D. and Black,M. (1985) Seeds, Physiology of Development and Germination. Plenum Press, New York, NY.
-
- Bryan J. and Aguilar-Bryan,L. (1999) Sulfonyl urea receptors: ABC transporters that regulate ATP-sensitive K+ channels. Biochim. Biophys. Acta, 1461, 285–303. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
