

Homeodomain protein ATHB6 is a target of the protein phosphatase ABI1 and regulates hormone responses in Arabidopsis
- PMID: 12065416
- PMCID: PMC126069
- DOI: 10.1093/emboj/cdf316
Homeodomain protein ATHB6 is a target of the protein phosphatase ABI1 and regulates hormone responses in Arabidopsis
Abstract
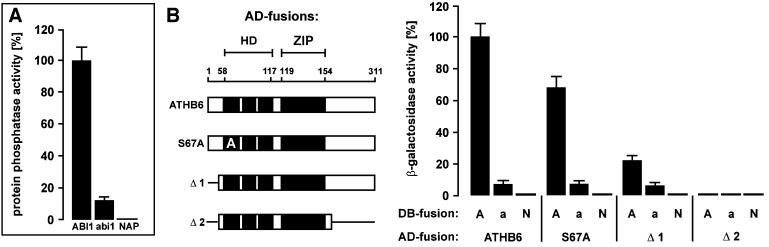
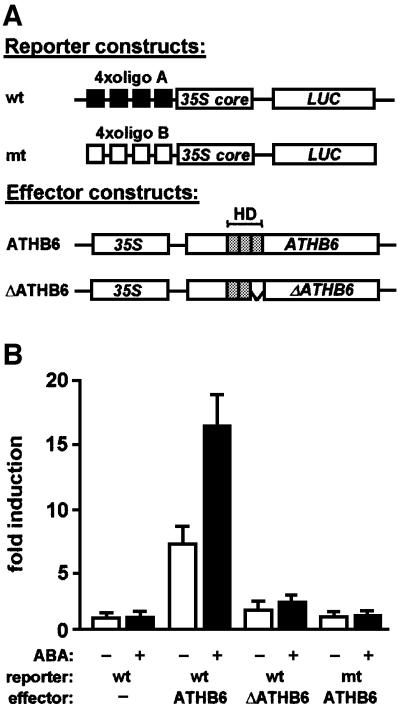
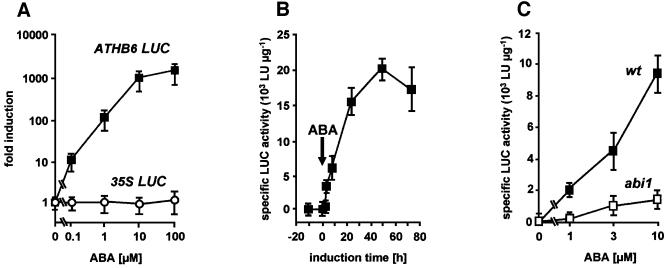
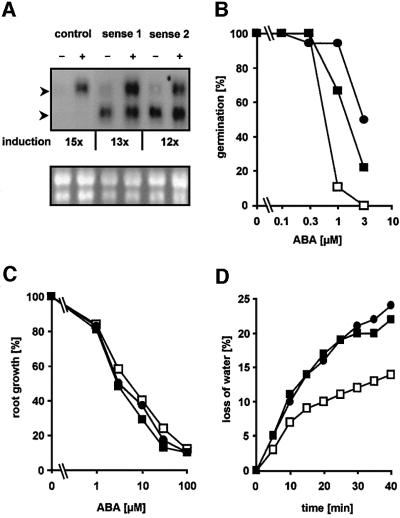
ABI1, a protein phosphatase 2C, is a key component of signal transduction in Arabidopsis. It regulates diverse responses to the phytohormone abscisic acid (ABA) such as stomatal closure, seed dormancy and inhibition of vegetative growth. By analysing proteins capable of interacting with ABI1, we have identified the homeodomain protein ATHB6 as a regulator of the ABA signal pathway. Critical for interaction between ATHB6 and ABI1 is an intact protein phosphatase domain and the N-terminal domain of ATHB6 containing the DNA-binding site. ATHB6 recognizes a cis-element present in its promoter, which encompasses the core motif (CAATTATTA) that mediated ATHB6- and ABA-dependent gene expression in protoplasts. In addition, transgenic plants containing a luciferase gene controlled by the ATHB6 promoter documented a strong ABA-inducible expression of the reporter which was abrogated in the ABA-insensitive abi1 mutant. Arabidopsis plants with constitutive expression of the transcriptional regulator revealed ABA insensitivity in a subset of ABI1-dependent responses. Thus, the homeodomain protein ATHB6 seems to represent a negative regulator of the ABA signal pathway and to act downstream of ABI1.
Figures


Similar articles
-
The HD-Zip gene ATHB6 in Arabidopsis is expressed in developing leaves, roots and carpels and up-regulated by water deficit conditions.Plant Mol Biol. 1999 Aug;40(6):1073-83. doi: 10.1023/a:1006267013170. Plant Mol Biol. 1999. PMID: 10527431
-
Nuclear localization of the mutant protein phosphatase abi1 is required for insensitivity towards ABA responses in Arabidopsis.Plant J. 2008 Jun;54(5):806-19. doi: 10.1111/j.1365-313X.2008.03454.x. Epub 2008 Feb 22. Plant J. 2008. PMID: 18298671
-
The regulatory domain of SRK2E/OST1/SnRK2.6 interacts with ABI1 and integrates abscisic acid (ABA) and osmotic stress signals controlling stomatal closure in Arabidopsis.J Biol Chem. 2006 Feb 24;281(8):5310-8. doi: 10.1074/jbc.M509820200. Epub 2005 Dec 19. J Biol Chem. 2006. PMID: 16365038
-
Regulation of Arabidopsis MAPKKK18 by ABI1 and SnRK2, components of the ABA signaling pathway.Plant Signal Behav. 2016;11(4):e1139277. doi: 10.1080/15592324.2016.1139277. Plant Signal Behav. 2016. PMID: 26852793 Free PMC article. Review.
-
Protein phosphatase 2C (PP2C) function in higher plants.Plant Mol Biol. 1998 Dec;38(6):919-27. doi: 10.1023/a:1006054607850. Plant Mol Biol. 1998. PMID: 9869399 Review.
Cited by
-
ABA inducible rice protein phosphatase 2C confers ABA insensitivity and abiotic stress tolerance in Arabidopsis.PLoS One. 2015 Apr 17;10(4):e0125168. doi: 10.1371/journal.pone.0125168. eCollection 2015. PLoS One. 2015. PMID: 25886365 Free PMC article.
-
Phospholipase dalpha1 and phosphatidic acid regulate NADPH oxidase activity and production of reactive oxygen species in ABA-mediated stomatal closure in Arabidopsis.Plant Cell. 2009 Aug;21(8):2357-77. doi: 10.1105/tpc.108.062992. Epub 2009 Aug 18. Plant Cell. 2009. PMID: 19690149 Free PMC article.
-
Enhancement of abscisic acid sensitivity and reduction of water consumption in Arabidopsis by combined inactivation of the protein phosphatases type 2C ABI1 and HAB1.Plant Physiol. 2006 Aug;141(4):1389-99. doi: 10.1104/pp.106.081018. Epub 2006 Jun 23. Plant Physiol. 2006. PMID: 16798945 Free PMC article.
-
Protein phosphatases 2C regulate the activation of the Snf1-related kinase OST1 by abscisic acid in Arabidopsis.Plant Cell. 2009 Oct;21(10):3170-84. doi: 10.1105/tpc.109.069179. Epub 2009 Oct 23. Plant Cell. 2009. PMID: 19855047 Free PMC article.
-
Functional analyses of the ABI1-related protein phosphatase type 2C reveal evolutionarily conserved regulation of abscisic acid signaling between Arabidopsis and the moss Physcomitrella patens.Plant Mol Biol. 2009 Jun;70(3):327-40. doi: 10.1007/s11103-009-9476-z. Epub 2009 Mar 6. Plant Mol Biol. 2009. PMID: 19266168
References
-
- Abel S. and Theologis,A. (1994) Transient transformation of Arabidopsis leaf protoplasts: a versatile experimental system to study gene expression. Plant J., 5, 421–427. - PubMed
-
- Allen G.J., Chu,S., Harrington,C.L., Schumacher,K., Hoffmann,T., Tang,Y.Y., Grill,E. and Schroeder,J.I. (2001) A defined range of guard cell calcium oscillation parameters encodes stomatal movements. Nature, 411, 1053–1057. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
