

R-loop-dependent rolling-circle replication and a new model for DNA concatemer resolution by mitochondrial plasmid mp1
- PMID: 12065425
- PMCID: PMC126065
- DOI: 10.1093/emboj/cdf311
R-loop-dependent rolling-circle replication and a new model for DNA concatemer resolution by mitochondrial plasmid mp1
Abstract
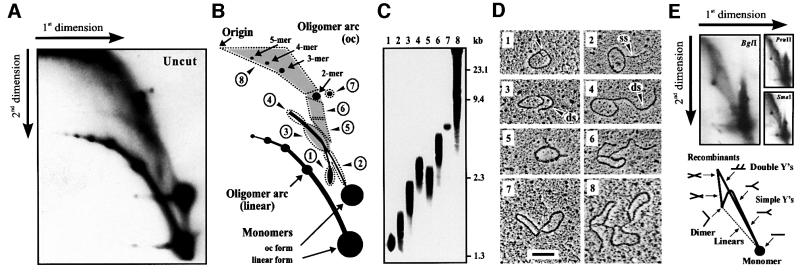
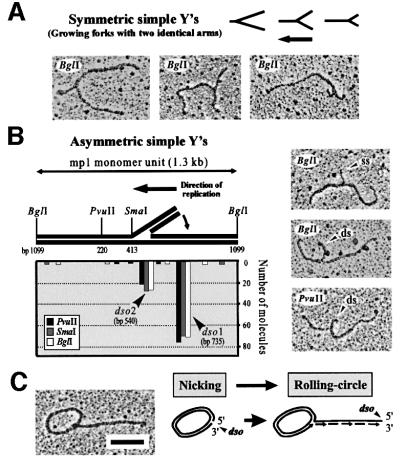
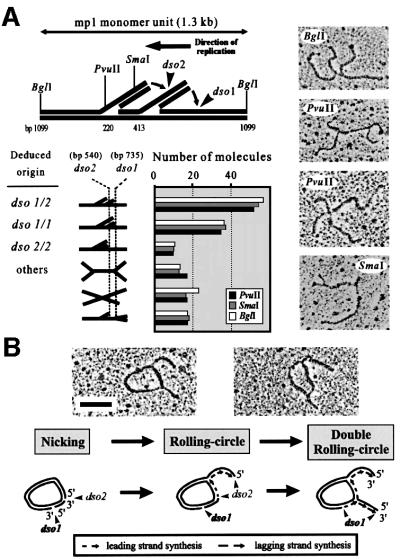
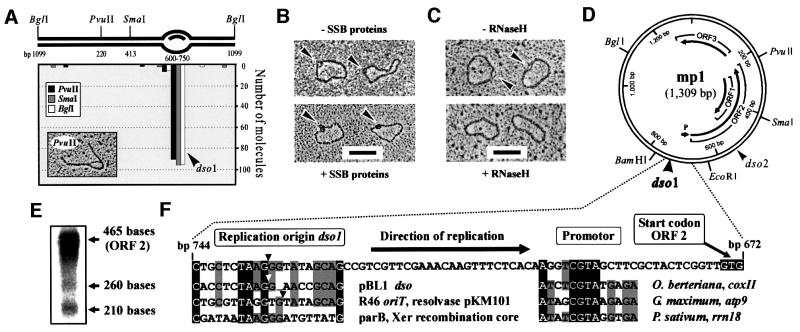
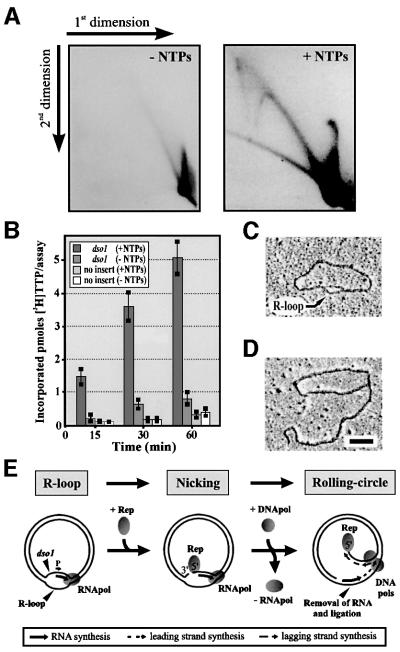
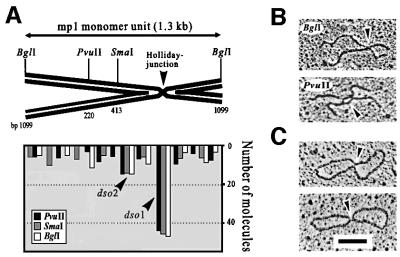
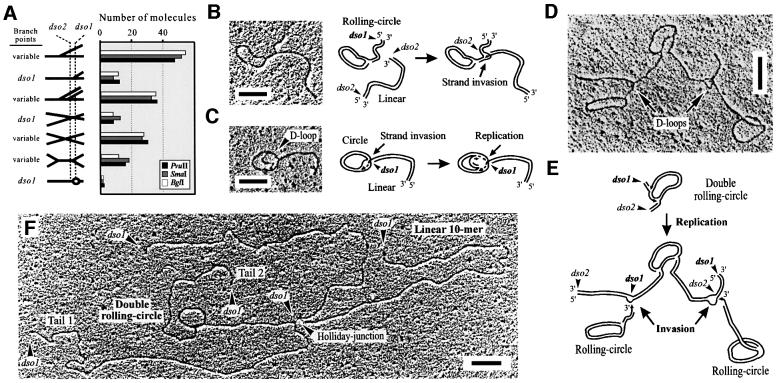
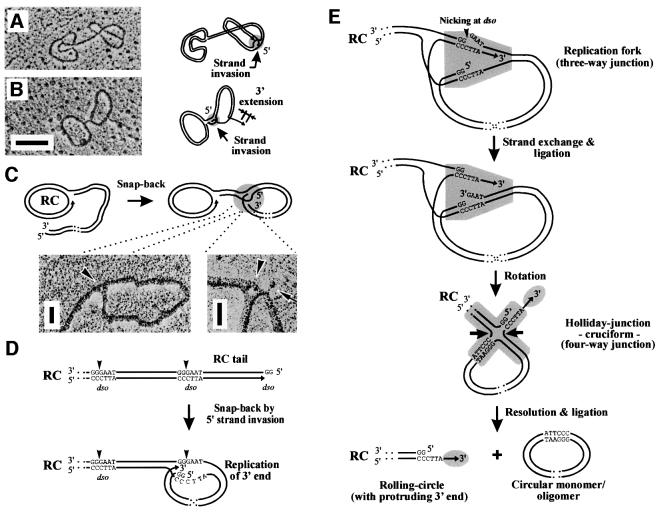
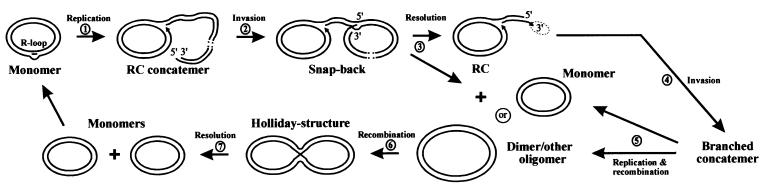
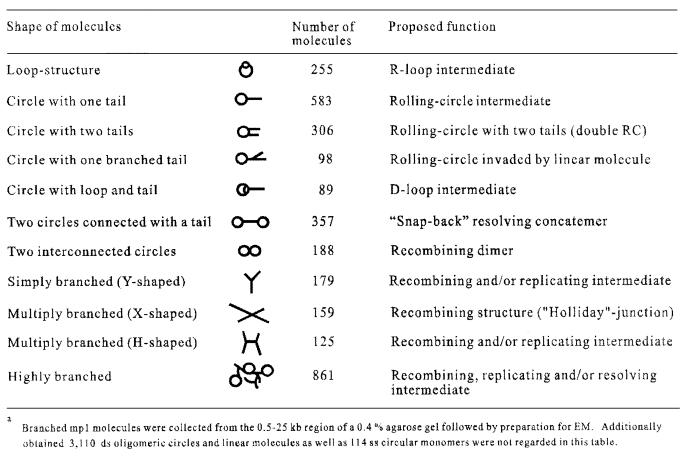
The mitochondrial (mt) plasmid mp1 of Chenopodium album replicates by a rolling-circle (RC) mechanism initiated at two double-stranded replication origins (dso1 and dso2). Two-dimensional gel electrophoresis and electron microscopy of early mp1 replication intermediates revealed novel spots. Ribonucleotide (R)-loops were identified at dso1, which function as a precursor for the RCs in vivo and in vitro. Bacteriophage T4-like networks of highly branched mp1 concatemers with up to 20 monomer units were mapped and shown to be mainly formed by replicating, invading, recombining and resolving molecules. A new model is proposed in which concatemers were separated into single units by a "snap-back" mechanism and homologous recombination. dso1 is a recombination hotspot, with sequence homology to bacterial Xer recombination cores. mp1 is a unique eukaryotic plasmid that expresses features of phages like T4 and could serve as a model system for replication and maintenance of DNA concatemers.
Figures
Similar articles
-
Rolling-Circle Replication in Mitochondrial DNA Inheritance: Scientific Evidence and Significance from Yeast to Human Cells.Genes (Basel). 2020 May 6;11(5):514. doi: 10.3390/genes11050514. Genes (Basel). 2020. PMID: 32384722 Free PMC article. Review.
-
In vitro replication of mitochondrial plasmid mp1 from the higher plant Chenopodium album (L.): a remnant of bacterial rolling circle and conjugative plasmids?J Mol Biol. 1998 Dec 11;284(4):1005-15. doi: 10.1006/jmbi.1998.2254. J Mol Biol. 1998. PMID: 9837722
-
Strand switching during rolling circle replication of plasmid-like DNA circles in the mitochondria of the higher plant Chenopodium album (L.).Plasmid. 2000 Mar;43(2):166-70. doi: 10.1006/plas.1999.1437. Plasmid. 2000. PMID: 10686137
-
Unique features of the mitochondrial rolling circle-plasmid mp1 from the higher plant Chenopodium album (L.).Nucleic Acids Res. 1997 Feb 1;25(3):582-89. doi: 10.1093/nar/25.3.582. Nucleic Acids Res. 1997. PMID: 9016599 Free PMC article.
-
Plasmid rolling-circle replication: highlights of two decades of research.Plasmid. 2005 Mar;53(2):126-36. doi: 10.1016/j.plasmid.2004.12.008. Epub 2005 Jan 22. Plasmid. 2005. PMID: 15737400 Review.
Cited by
-
What we know so far and what we can expect next: A molecular investigation of plant parasitism.Genet Mol Biol. 2024 Sep 30;47Suppl 1(Suppl 1):e20240051. doi: 10.1590/1678-4685-GMB-2024-0051. eCollection 2024. Genet Mol Biol. 2024. PMID: 39348487 Free PMC article.
-
Recombination at long mutant telomeres produces tiny single- and double-stranded telomeric circles.Mol Cell Biol. 2005 Jun;25(11):4406-12. doi: 10.1128/MCB.25.11.4406-4412.2005. Mol Cell Biol. 2005. PMID: 15899847 Free PMC article.
-
Rolling-Circle Replication in Mitochondrial DNA Inheritance: Scientific Evidence and Significance from Yeast to Human Cells.Genes (Basel). 2020 May 6;11(5):514. doi: 10.3390/genes11050514. Genes (Basel). 2020. PMID: 32384722 Free PMC article. Review.
-
Mhr1p-dependent concatemeric mitochondrial DNA formation for generating yeast mitochondrial homoplasmic cells.Mol Biol Cell. 2004 Jan;15(1):310-22. doi: 10.1091/mbc.e03-07-0508. Epub 2003 Oct 17. Mol Biol Cell. 2004. PMID: 14565971 Free PMC article.
-
Double-strand break repair processes drive evolution of the mitochondrial genome in Arabidopsis.BMC Biol. 2011 Sep 27;9:64. doi: 10.1186/1741-7007-9-64. BMC Biol. 2011. PMID: 21951689 Free PMC article.
References
-
- Backert S. (2000) Strand switching during rolling circle replication of plasmid-like DNA circles in the mitochondria of the higher plant Chenopodium album (L.). Plasmid, 43, 166–170. - PubMed
-
- Backert S., Kunnimalaiyaan,M., Börner,T. and Nielsen,B.L. (1998) In vitro replication of mitochondrial plasmid mp1 from the higher plant Chenopodium album (L.): a remnant of bacterial rolling circle and conjugative plasmids? J. Mol. Biol., 284, 1005–1015. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
