

Drf1, a novel regulatory subunit for human Cdc7 kinase
- PMID: 12065429
- PMCID: PMC126049
- DOI: 10.1093/emboj/cdf290
Drf1, a novel regulatory subunit for human Cdc7 kinase
Abstract
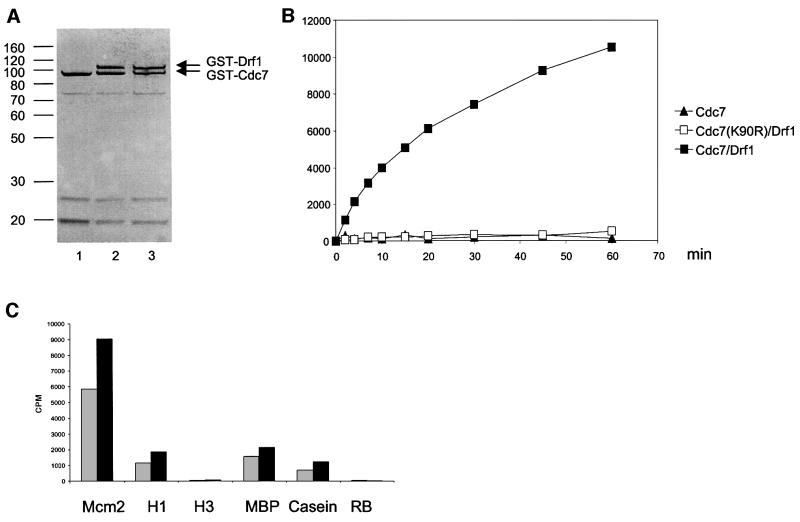
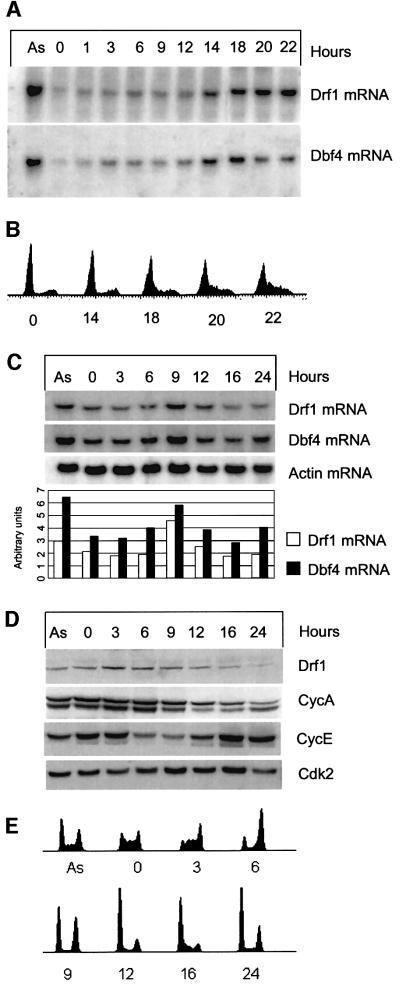
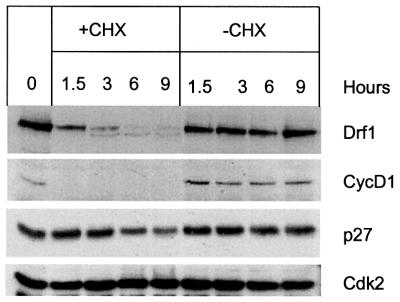
Studies in model organisms have contributed to elucidate multiple levels at which regulation of eukaryotic DNA replication occurs. Cdc7, an evolutionarily conserved serine-threonine kinase, plays a pivotal role in linking cell cycle regulation to genome duplication, being essential for the firing of DNA replication origins. Binding of the cell cycle-regulated subunit Dbf4 to Cdc7 is necessary for in vitro kinase activity. This binding is also thought to be the key regulatory event that controls Cdc7 activity in cells. Here, we describe a novel human protein, Drf1, related to both human and yeast Dbf4. Drf1 is a nuclear cell cycle-regulated protein, it binds to Cdc7 and activates the kinase. Therefore, human Cdc7, like cyclin-dependent kinases, can be activated by alternative regulatory subunits. Since the Drf1 gene is either absent or not yet identified in the genome of model organisms such as yeast and Drosophila, these findings introduce a new level of complexity in the regulation of DNA replication of the human genome.
Figures



References
-
- Brown G.W. and Kelly,T.J. (1998) Purification of Hsk1, a minichromosome maintenance protein kinase from fission yeast. J. Biol. Chem., 273, 22083–22090. - PubMed
-
- Diffley J.F. and Labib,K. (2002) The chromosome replication cycle. J. Cell Sci., 115, 869–872. - PubMed
-
- Dimitrova D.S. and Gilbert,D.M. (1999) The spatial position and replication timing of chromosomal domains are both established in early G1 phase. Mol. Cell, 4, 983–993. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
