

Carrier-mediated uptake and release of taurine from Bergmann glia in rat cerebellar slices
- PMID: 12068038
- PMCID: PMC2290349
- DOI: 10.1113/jphysiol.2001.015834
Carrier-mediated uptake and release of taurine from Bergmann glia in rat cerebellar slices
Abstract
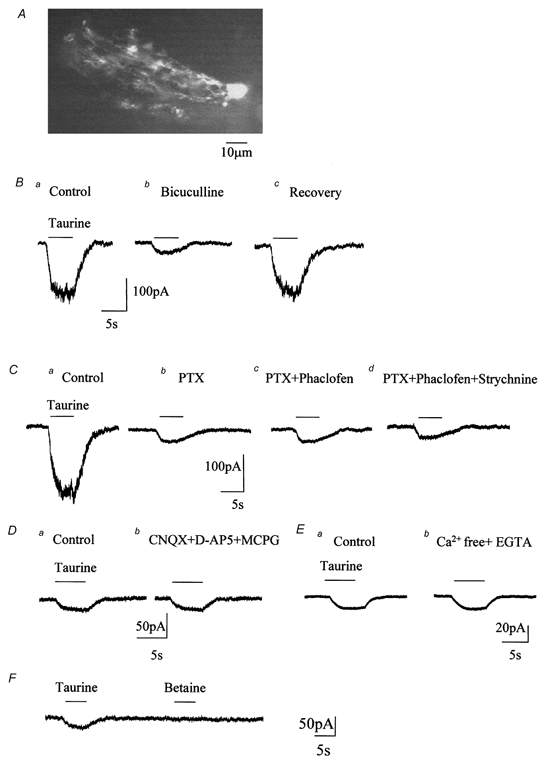
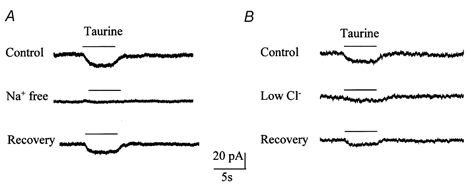
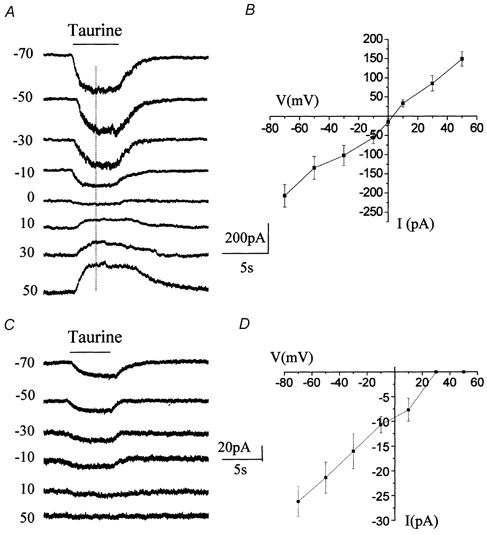
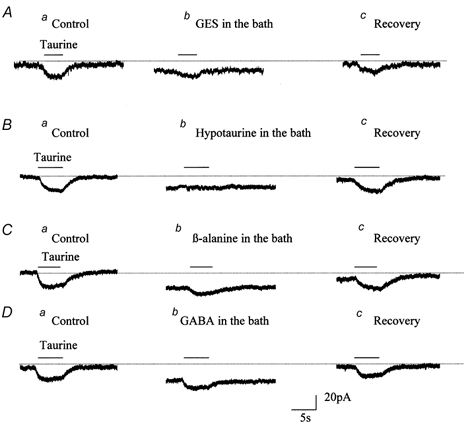
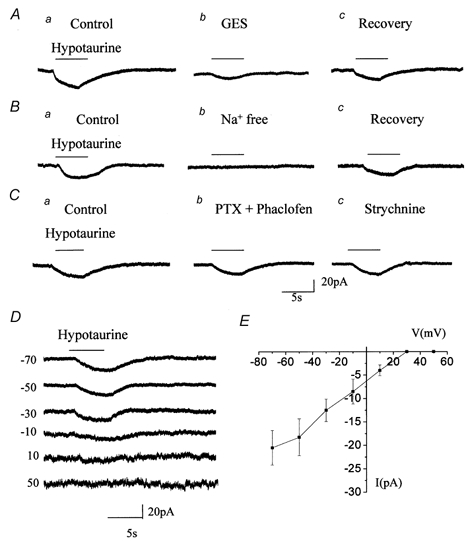
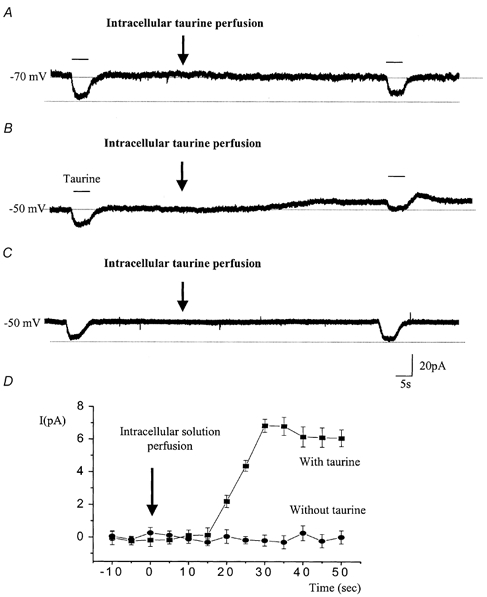
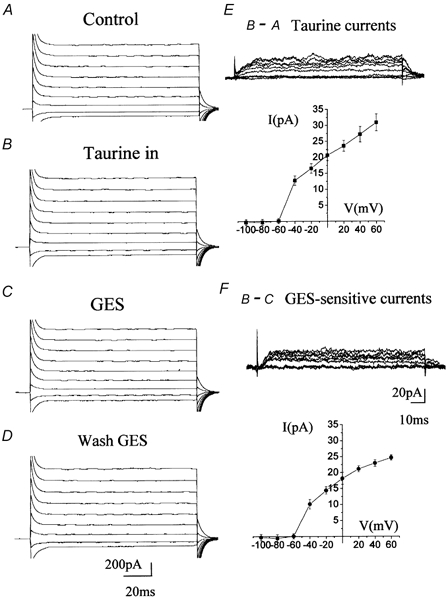
Taurine uptake is essential for the maintenance of millimolar intracellular concentrations of taurine, which is released during ischaemia and is thought to be neuroprotective. To determine whether Bergmann glia express functional transporters that can mediate both taurine uptake and efflux, whole-cell patch-clamp recordings were obtained from these cells in rat cerebellar slices. Taurine-induced inward currents can be pharmacologically separated into GABA(A) receptor and taurine transporter currents. In the presence of GABA receptor blockers, residual taurine currents averaged -28 pA at -70 mV and were strictly inwardly rectifying between -70 and +50 mV. These residual currents were also abolished by external Na+ removal and diminished by reduction of external Cl-, consistent with transport currents. Taurine transport currents were reduced by a taurine transporter inhibitor, guanidinoethyl sulphonate (GES). Other classical inhibitors reduced taurine transport currents with an order of potency (hypotaurine > beta-alanine > GES > GABA) similar to that reported for cloned rat taurine transporters. Following intracellular taurine perfusion during the recording, a progressively developing outward current could be observed at -50 mV but not at -70 mV. Intracellular perfusion of taurine also decreased taurine-induced inward currents at both holding potentials. Outward currents induced by intracellular taurine increased in amplitude with depolarization, activated near -50 mV, and were affected by GES. For the first time, these results demonstrate that taurine activates both GABA(A) receptors and Na+/Cl--dependent taurine transporters in Bergmann glia in slices. In addition, our data show that taurine transporters can work in reverse and can probably mediate taurine efflux under ischaemic conditions.
Figures
References
-
- Albrecht J, Bender AS, Norenberg MD. Ammonia stimulates the release of taurine from cultured astrocytes. Brain Research. 1994;660:288–292. - PubMed
-
- Almarghini K, Remy A, Tappaz M. Immunocyto-chemistry of the taurine biosynthesis enzyme, cysteine sulfinate decarboxylase, in the cerebellum: evidence for a glial localization. Neuroscience. 1991;43:111–119. - PubMed
-
- Andjelkovic AV, Kerkovich D, Shanley J, Pulliam L, Pachter JS. Expression of binding sites for β chemokines on human astrocytes. Glia. 1999;28:225–235. - PubMed
-
- Attwell D, Barbour B, Szatkowski M. Non-vesicular release of neurotransmitter. Neuron. 1993;11:401–407. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources