

Interleukin 15 is required for proliferative renewal of virus-specific memory CD8 T cells
- PMID: 12070282
- PMCID: PMC2193552
- DOI: 10.1084/jem.20020369
Interleukin 15 is required for proliferative renewal of virus-specific memory CD8 T cells
Abstract
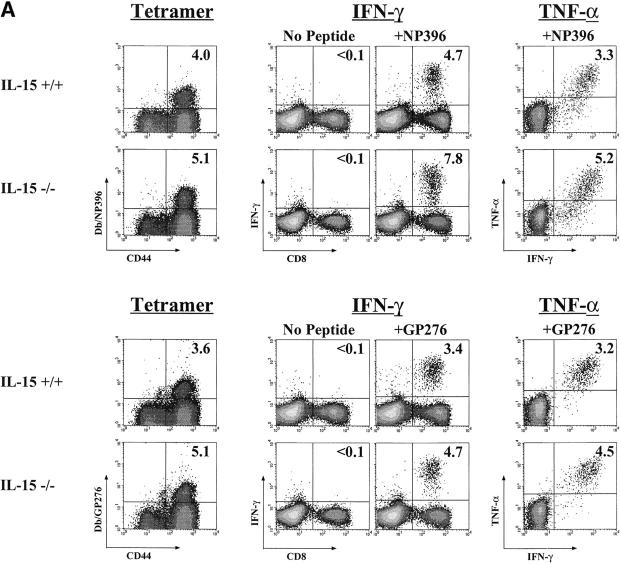
The generation and efficient maintenance of antigen-specific memory T cells is essential for long-lasting immunological protection. In this study, we examined the role of interleukin (IL)-15 in the generation and maintenance of virus-specific memory CD8 T cells using mice deficient in either IL-15 or the IL-15 receptor alpha chain. Both cytokine- and receptor-deficient mice made potent primary CD8 T cell responses to infection with lymphocytic choriomeningitis virus (LCMV), effectively cleared the virus and generated a pool of antigen-specific memory CD8 T cells that were phenotypically and functionally similar to memory CD8 T cells present in IL-15(+/+) mice. However, longitudinal analysis revealed a slow attrition of virus-specific memory CD8 T cells in the absence of IL-15 signals. This loss of CD8 T cells was due to a severe defect in the proliferative renewal of antigen-specific memory CD8 T cells in IL-15(-/-) mice. Taken together, these results show that IL-15 is not essential for the generation of memory CD8 T cells, but is required for homeostatic proliferation to maintain populations of memory cells over long periods of time.
Figures









Comment in
-
Multiple choices: regulation of memory CD8 T cell generation and homeostasis by interleukin (IL)-7 and IL-15.J Exp Med. 2002 Jun 17;195(12):F49-52. doi: 10.1084/jem.20020767. J Exp Med. 2002. PMID: 12070294 Free PMC article. No abstract available.
References
-
- Ahmed, R., and D. Gray. 1996. Immunological memory and protective immunity: understanding their relation. Science. 272:54–60. - PubMed
-
- Bachmann, M.F., M. Barner, A. Viola, and M. Kopf. 1999. Distinct kinetics of cytokine production and cytolysis in effector and memory T cells after viral infection. Eur. J. Immunol. 29:291–299. - PubMed
-
- Veiga-Fernandes, H., U. Walter, C. Bourgeois, A. McLean, and B. Rocha. 2000. Response of naive and memory CD8+ T cells to antigen stimulation in vivo. Nat. Immunol. 1:47–53. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
