

Estrogen alters thresholds for B cell apoptosis and activation
- PMID: 12070310
- PMCID: PMC151010
- DOI: 10.1172/JCI14873
Estrogen alters thresholds for B cell apoptosis and activation
Abstract
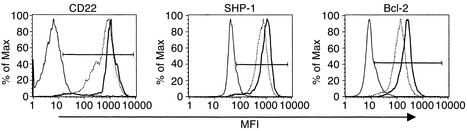
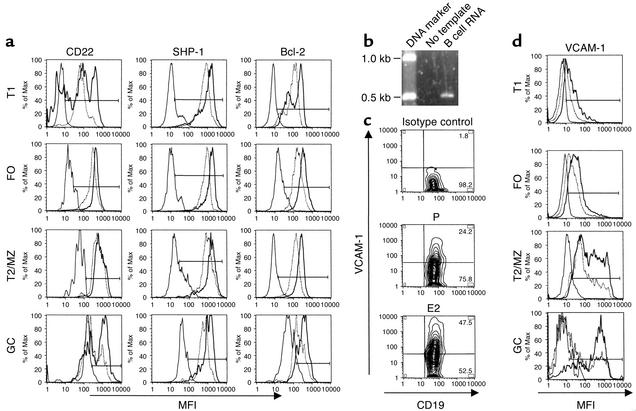
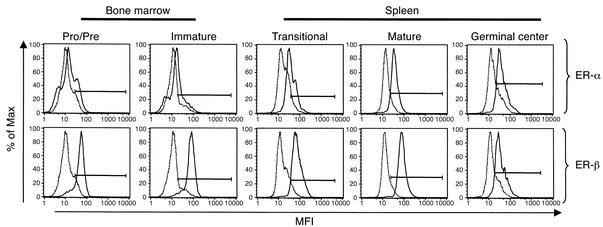
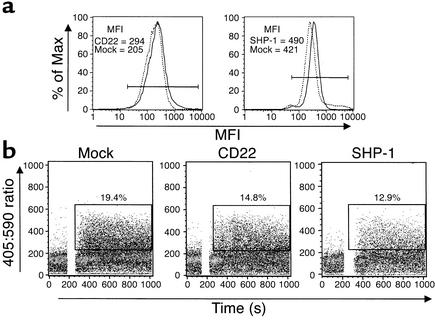
Estrogen is thought to contribute to the increased frequency of autoimmune disorders occurring in females, but a molecular basis for its effects on autoimmunity remains to be elucidated. We have shown previously that estrogen leads to the survival and activation of autoreactive cells in the naive repertoire. To identify the molecular pathways involved in B cell tolerance, we sought to identify genes that are differentially regulated by estrogen in mouse B cells. Several genes involved in B cell activation and survival, including cd22, shp-1, bcl-2, and vcam-1, were upregulated by estrogen in B cells. We found that overexpression of CD22 and SHP-1 in B cells decreased B cell receptor signaling. Estrogen receptors alpha and beta are expressed on B cells and are functional, since they can directly upregulate expression of CD22, SHP-1, and Bcl-2. Estrogen treatment protected isolated primary B cells from B cell receptor-mediated apoptosis. These results suggest that estrogen induces a genetic program that alters survival and activation of B cells in a B cell-autonomous fashion and thus skews the naive immune system toward autoreactivity.
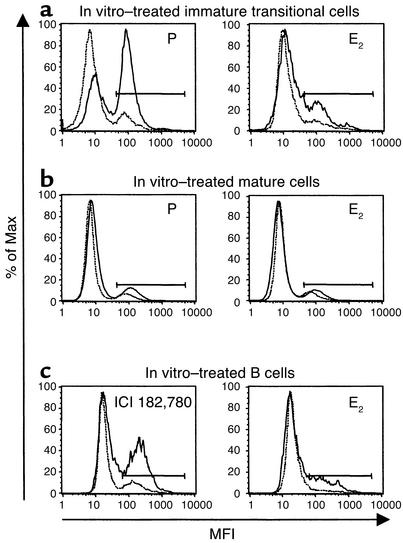
Figures
References
-
- Smithson G, Couse JF, Lubahn DB, Korach KS, Kincade PW. The role of estrogen receptors and androgen receptors in sex steroid regulation of B lymphopoiesis. J Immunol. 1998;161:27–34. - PubMed
-
- Butterworth M, McClellan B, Allansmith M. Influence of sex in immunoglobulin levels. Nature. 1967;214:1224–1225. - PubMed
-
- Schuurs AH, Verheul HA. Effects of gender and sex steroids on the immune response. J Steroid Biochem. 1990;35:157–172. - PubMed
-
- Grossman CJ, Roselle GA, Mendenhall CL. Sex steroid regulation of autoimmunity. J Steroid Biochem Mol Biol. 1991;40:649–659. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
