

ATP as a putative sensory mediator: activation of intrinsic sensory neurons of the myenteric plexus via P2X receptors
- PMID: 12077173
- PMCID: PMC6757757
- DOI: 10.1523/JNEUROSCI.22-12-04767.2002
ATP as a putative sensory mediator: activation of intrinsic sensory neurons of the myenteric plexus via P2X receptors
Abstract
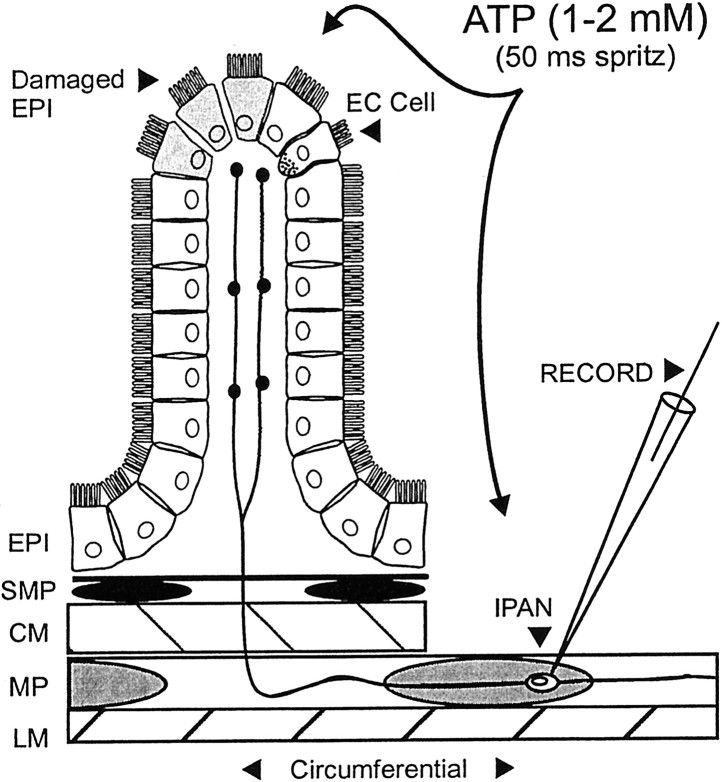
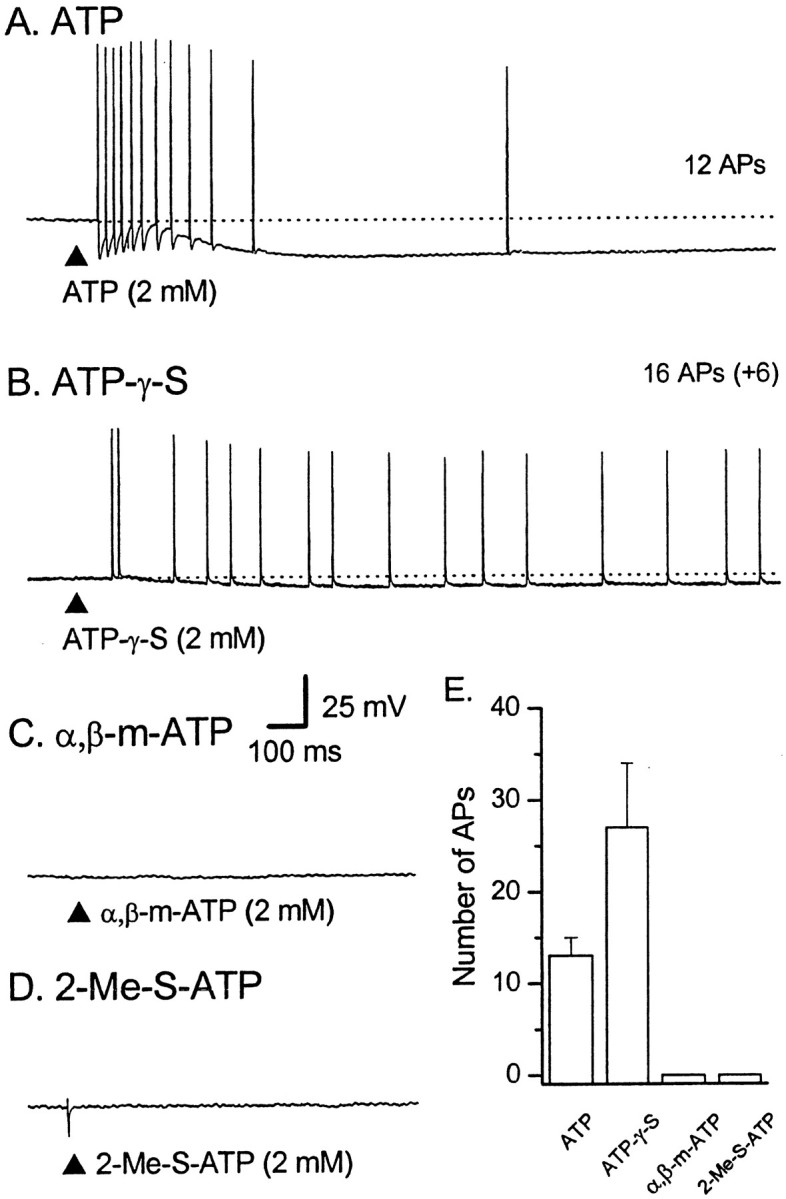
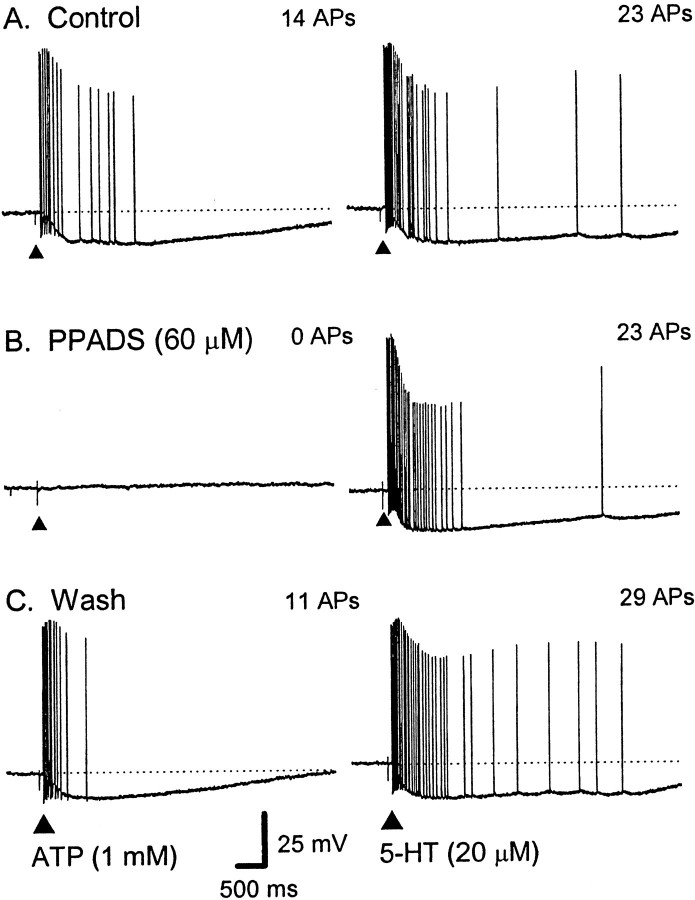
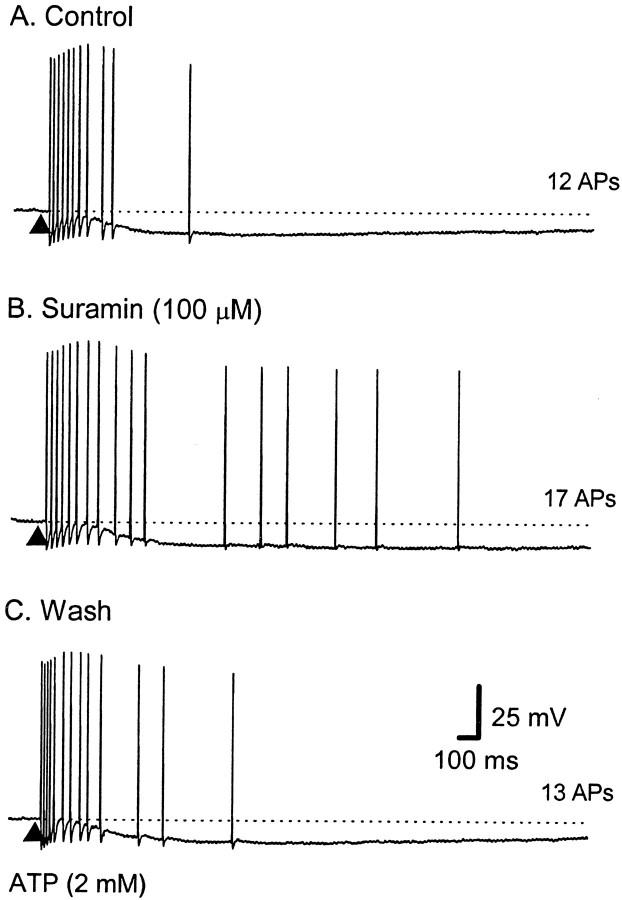
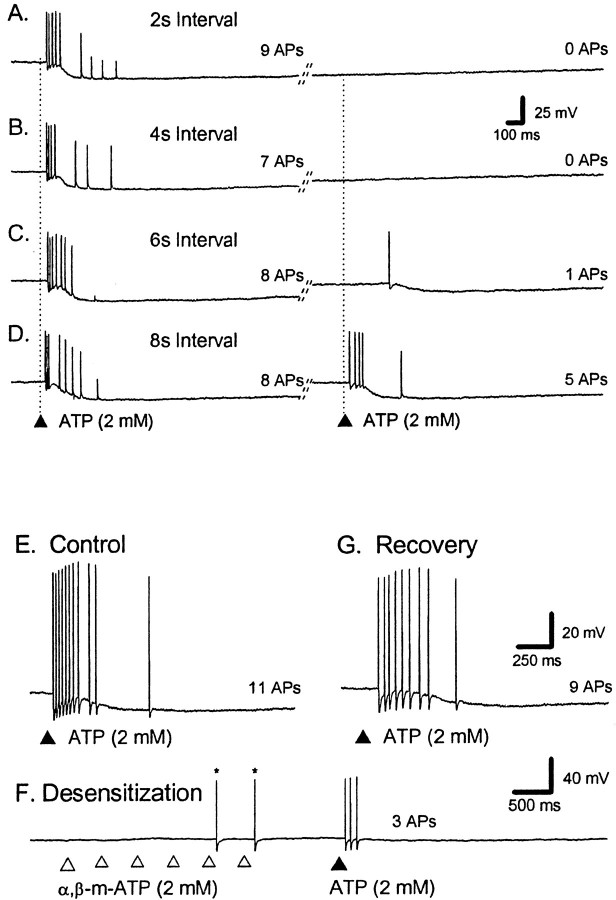
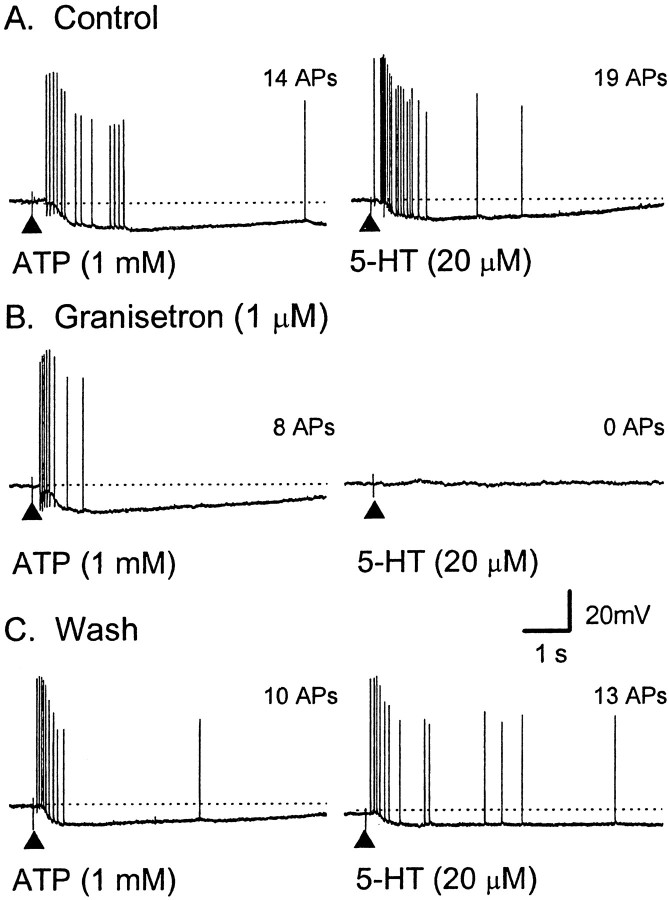
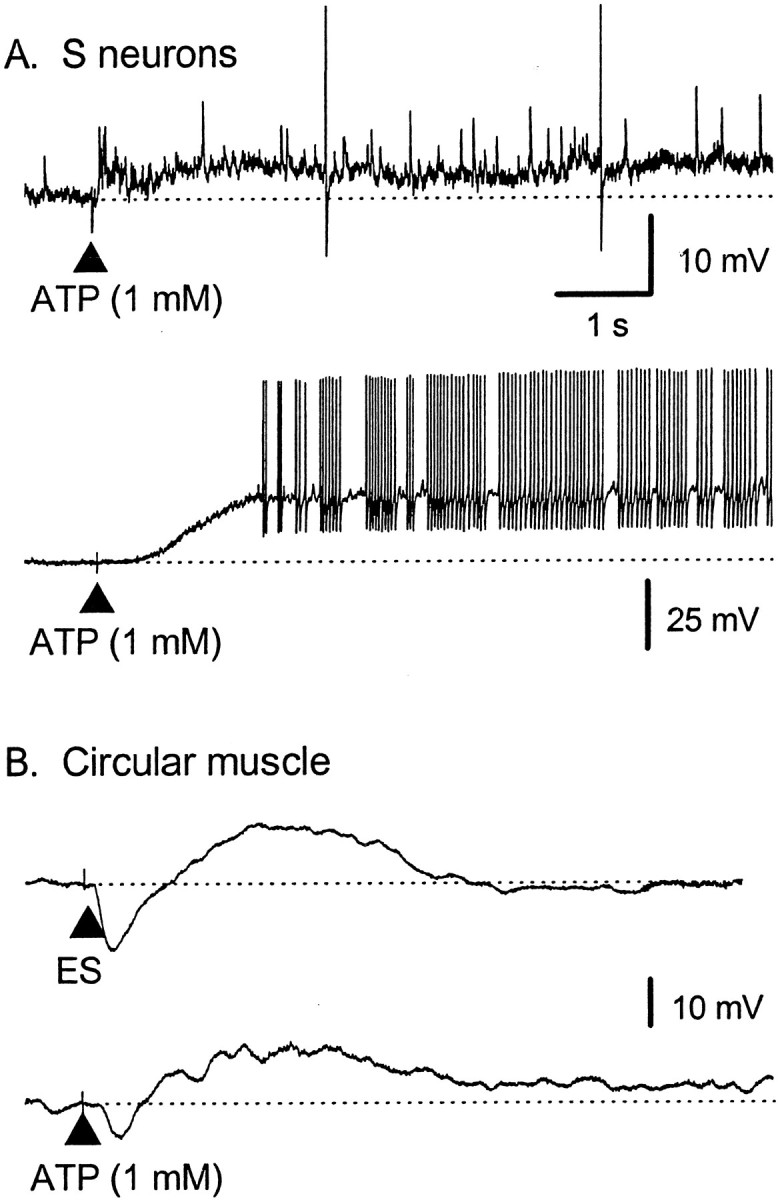
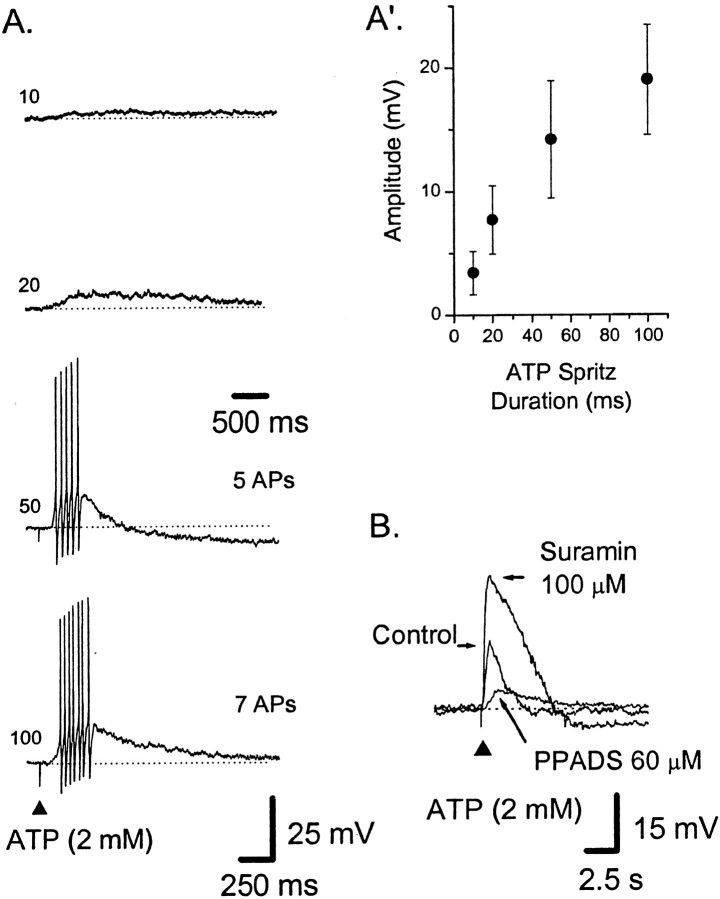
The mucosal terminals of sensory neurons intrinsic to the wall of the intestine are sensitive to the chemical environment within the lumen. Lumenal stimuli probably release sensory mediators from the mucosal epithelium, which then activate the nerve terminals indirectly. Here, we tested the idea that ATP activates intrinsic sensory nerve terminals in a way consistent with its being a sensory mediator. We made intracellular recordings from intrinsic sensory neurons located in the myenteric plexus [identified as AH neurons, which are neurons with a long-lasting afterhyperpolarization following the action potential (AP)], located within 1 mm of intact mucosa. Focal electrical stimulation of the mucosa was used to locate and map regions innervated by each neuron. Application of ATP (1-2 mm in the pressure pipette) to these regions elicited trains of APs that originated at the sensory terminals. ATP-gamma-S produced a similar response, but alpha,beta-methylene ATP and 2-methylthio-ATP were only weakly active. The P2 receptor antagonist pyridoxalphosphate-6-azophenyl-2',5'-disulphonic acid (PPADS) (60 microm in the bath) abolished the APs evoked by ATP and ATP-gamma-S but spared similar responses evoked by 5-hydroxytryptamine (5-HT). Another P2 receptor antagonist suramin (100 microm in the bath) did not significantly change the number of APs evoked by ATP. Either ATP or alpha,beta-methylene ATP desensitized the ATP-evoked APs; 50% recovery occurred after approximately 5 sec. The number of APs evoked by ATP was reduced, but not abolished, by the selective 5-HT3 receptor antagonist granisetron (1 microm in the bath). ATP was applied to the cell bodies of sensory neurons to investigate whether the cell bodies express the same P2X receptor as the terminals. ATP evoked a fast depolarization associated with a reduction in input resistance and a reversal potential of -11 mV. This depolarization was potentiated by suramin and blocked by PPADS. We conclude that activation of an atypical excitatory P2X receptor by ATP triggers AP generation in the mucosal processes of the sensory neurons; endogenous 5-HT release may also contribute to activation of the nerve terminals. A similar P2X receptor exists on the cell body of the sensory neuron. Together, these data are consistent with a role for ATP as a sensory mediator in gastrointestinal chemosensory transduction.
Figures
References
-
- Bertrand PP, Galligan JJ. Alfaxalone, pentobarbital and diazepam potentiate γ-aminobutyric acid-induced depolarizations in single myenteric neurons of guinea pig intestine. J Pharmacol Exp Ther. 1992;262:677–682. - PubMed
-
- Bertrand PP, Kunze WAA, Bornstein JC, Furness JB, Smith ML. Analysis of the responses of myenteric neurons in the small intestine to chemical stimulation of the mucosa. Am J Physiol. 1997;273:G422–G435. - PubMed
-
- Bertrand PP, Kunze WA, Bornstein JC, Furness JB. Electrical mapping of the projections of intrinsic primary afferent neurons to the mucosa of the guinea-pig small intestine. Neurogastroenterol Motil. 1998;10:533–541. - PubMed
-
- Bertrand PP, Kunze WAA, Furness JB, Bornstein JC. The terminals of myenteric intrinsic primary afferent neurons of the guinea-pig ileum are excited by 5-hydroxytryptamine acting at 5-hydroxytryptamine-3 receptors. Neuroscience. 2000;101:459–469. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous