

Yng1p modulates the activity of Sas3p as a component of the yeast NuA3 Hhistone acetyltransferase complex
- PMID: 12077334
- PMCID: PMC139787
- DOI: 10.1128/MCB.22.14.5047-5053.2002
Yng1p modulates the activity of Sas3p as a component of the yeast NuA3 Hhistone acetyltransferase complex
Abstract
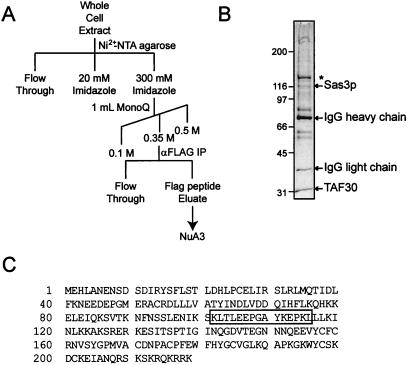
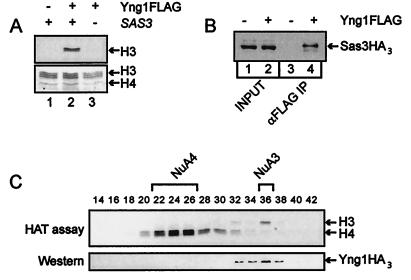
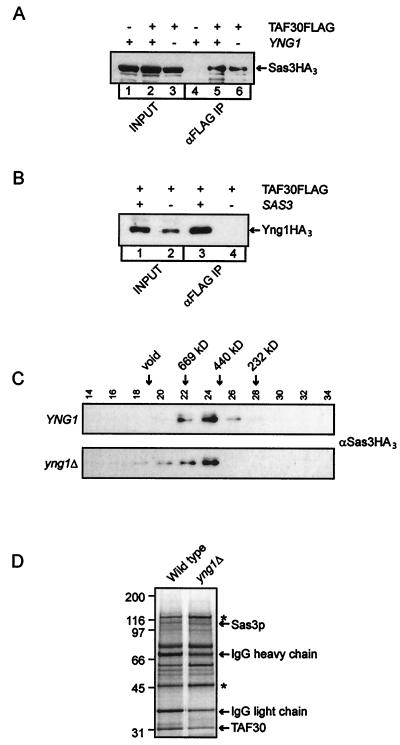
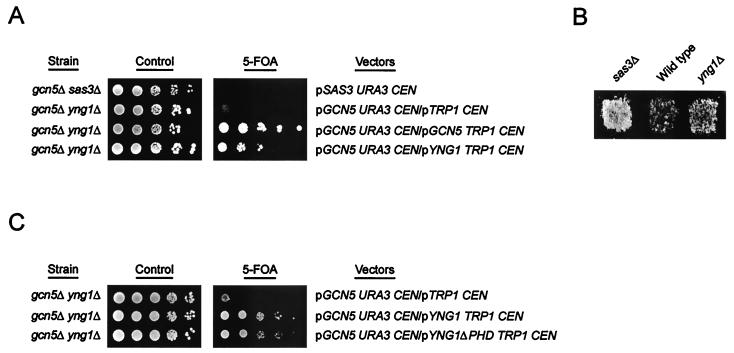
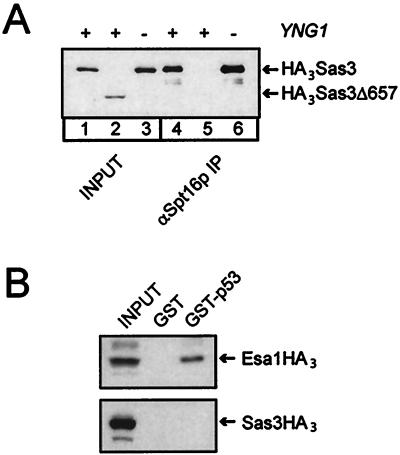
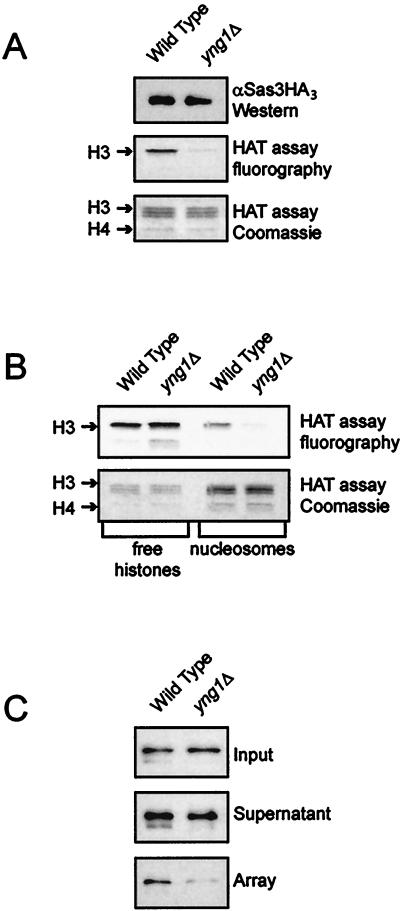
The mammalian ING1 gene encodes a tumor suppressor required for the function of p53. In this study we report a novel function for YNG1, a yeast homolog of ING1. Yng1p is a stable component of the NuA3 histone acetyltransferase complex, which contains Sas3p, the yeast homolog of the mammalian MOZ proto-oncogene product, as its catalytic subunit. Yng1p is required for NuA3 function in vivo but surprisingly is not required for the integrity of the complex. Instead, we find that Yng1p mediates the interaction of Sas3p with nucleosomes and is thus required for the ability of NuA3 to modify histone tails. These data, and the observations that other ING1 homologs are found in additional yeast complexes that posttranslationally modify histones, suggest that members of the ING1 class of proteins may have broad roles in enhancing or modifying the activities of chromatin-modifying complexes, thereby regulating their activities in transcription control.
Figures
References
-
- Aasland, R., T. J. Gibson, and A. F. Stewart. 1995. The PHD finger: implications for chromatin-mediated transcriptional regulation. Trends Biochem. Sci. 20:56-59. - PubMed
-
- Ausubel, F. M. 1987. Current protocols in molecular biology. J. Wiley, New York, N.Y.
-
- Balasubramanian, R., M. G. Pray-Grant, W. Selleck, P. A. Grant, and S. Tan. 2002. Role of the Ada2 and Ada3 transcriptional coactivators in histone acetylation. J. Biol. Chem. 277:7989-7995. - PubMed
-
- Brown, C. E., L. Howe, K. Sousa, S. C. Alley, M. J. Carrozza, S. Tan, and J. L. Workman. 2001. Recruitment of HAT complexes by direct activator interactions with the ATM-related Tra1 subunit. Science 292:2333-2337. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous