

FIH-1 is an asparaginyl hydroxylase enzyme that regulates the transcriptional activity of hypoxia-inducible factor
- PMID: 12080085
- PMCID: PMC186346
- DOI: 10.1101/gad.991402
FIH-1 is an asparaginyl hydroxylase enzyme that regulates the transcriptional activity of hypoxia-inducible factor
Abstract
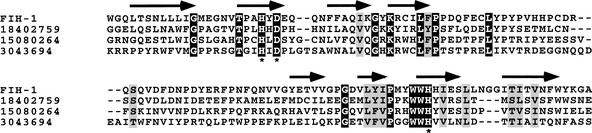
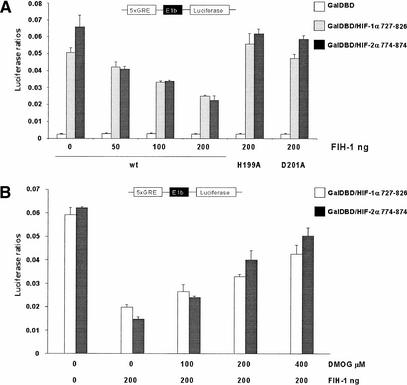
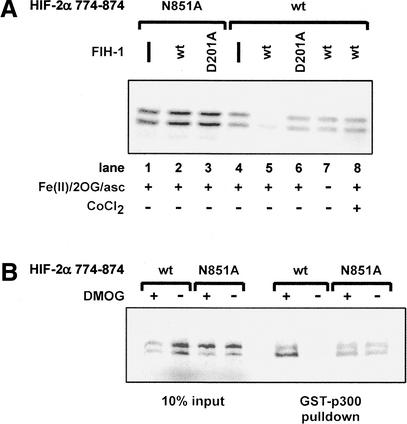
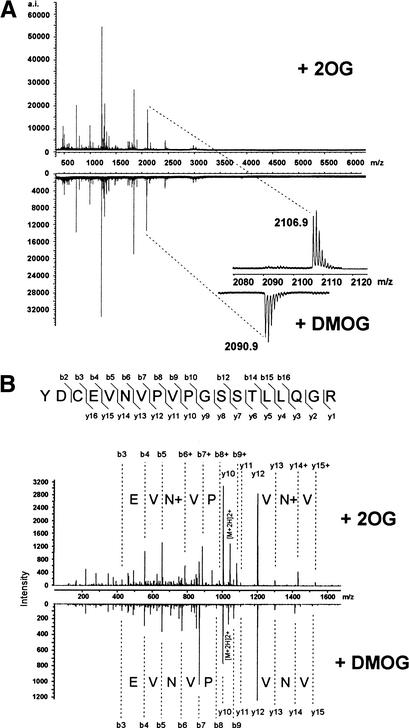
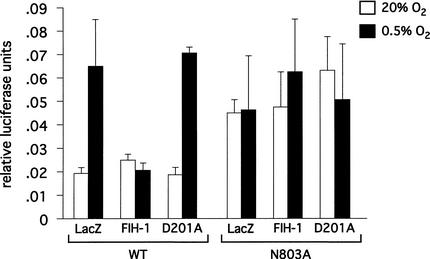
Mammalian cells adapt to hypoxic conditions through a transcriptional response pathway mediated by the hypoxia-inducible factor, HIF. HIF transcriptional activity is suppressed under normoxic conditions by hydroxylation of an asparagine residue within its C-terminal transactivation domain, blocking association with coactivators. Here we show that the protein FIH-1, previously shown to interact with HIF, is an asparaginyl hydroxylase. Like known hydroxylase enzymes, FIH-1 is an Fe(II)-dependent enzyme that uses molecular O(2) to modify its substrate. Together with the recently discovered prolyl hydroxylases that regulate HIF stability, this class of oxygen-dependent enzymes comprises critical regulatory components of the hypoxic response pathway.
Figures
References
-
- Bruick RK, McKnight SL. A conserved family of prolyl-4-hydroxylases that modify HIF. Science. 2001;294:1337–1340. - PubMed
-
- Clifton IJ, Hsueh L-C, Baldwin JE, Harlos K, Schofield CJ. Structure of proline 3-hydroxylase. Evolution of the family of 2-oxoglutarate dependent oxygenases. Eur J Biochem. 2001;268:6625–6636. - PubMed
-
- Cockman ME, Masson N, Mole DR, Jaakkola P, Chang GW, Clifford SC, Maher ER, Pugh CW, Ratcliffe PJ, Maxwell PH. Hypoxia inducible factor-α binding and ubiquitylation by the von Hippel-Lindau tumor suppressor protein. J Biol Chem. 2000;275:25733–25741. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases