

Inactivation of cytochrome o ubiquinol oxidase relieves catabolic repression of the Pseudomonas putida GPo1 alkane degradation pathway
- PMID: 12081947
- PMCID: PMC135178
- DOI: 10.1128/JB.184.14.3785-3793.2002
Inactivation of cytochrome o ubiquinol oxidase relieves catabolic repression of the Pseudomonas putida GPo1 alkane degradation pathway
Abstract
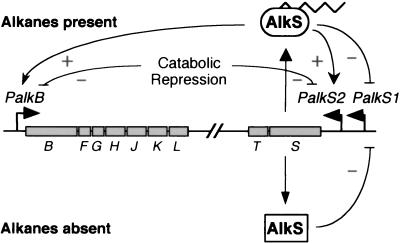
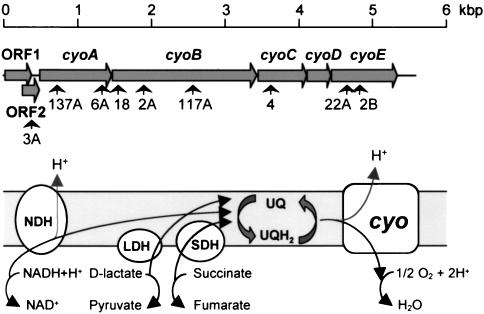
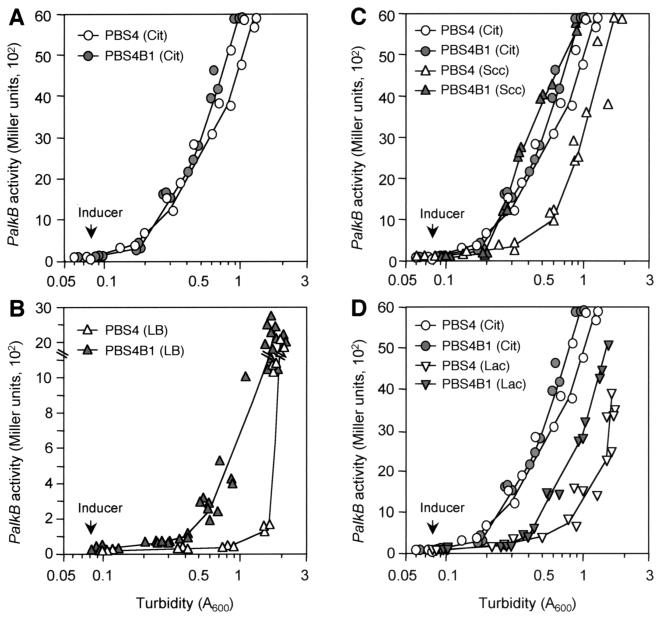
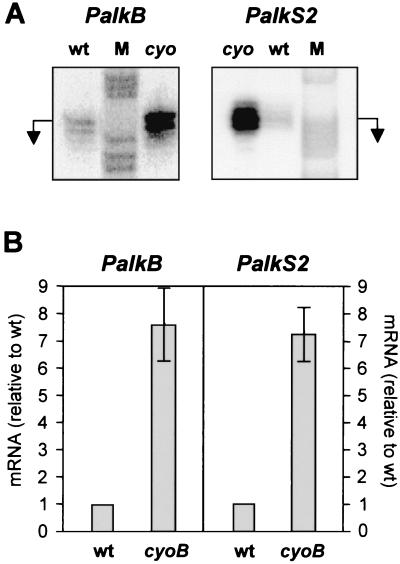
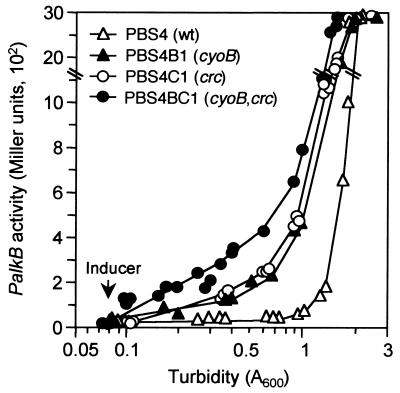
Expression of the alkane degradation pathway encoded by the OCT plasmid of Pseudomonas putida GPo1 is regulated by two control systems. One relies on the transcriptional regulator AlkS, which activates expression of the pathway in the presence of alkanes. The other, which is a dominant global regulation control, represses the expression of the pathway genes when a preferred carbon source is present in the growth medium in addition to alkanes. This catabolite repression control occurs through a poorly characterized mechanism that ultimately regulates transcription from the two AlkS-activated promoters of the pathway. To identify the factors involved, a screening method was developed to isolate mutants without this control. Several isolates were obtained, all of which contained mutations that mapped to genes encoding cytochrome o ubiquinol oxidase, the main terminal oxidase of the electron transport chain under highly aerobic conditions. Elimination of this terminal oxidase led to a decrease in the catabolic repression observed both in rich Luria-Bertani medium and in a defined medium containing lactate or succinate as the carbon source. This suggests that catabolic repression could monitor the physiological or metabolic status by using information from the electron transport chain or from the redox state of the cell. Since inactivation of the crc gene also reduces catabolic repression in rich medium (although not that observed in a defined medium), a strain was generated lacking both the Crc function and the cytochrome o terminal oxidase. The two mutations had an additive effect in relieving catabolic repression in rich medium. This suggests that crc and cyo belong to different regulation pathways, both contributing to catabolic repression.
Figures
Similar articles
-
Role of the crc gene in catabolic repression of the Pseudomonas putida GPo1 alkane degradation pathway.J Bacteriol. 2001 Nov;183(21):6197-206. doi: 10.1128/JB.183.21.6197-6206.2001. J Bacteriol. 2001. PMID: 11591662 Free PMC article.
-
Expression of the Pseudomonas putida OCT plasmid alkane degradation pathway is modulated by two different global control signals: evidence from continuous cultures.J Bacteriol. 2003 Aug;185(16):4772-8. doi: 10.1128/JB.185.16.4772-4778.2003. J Bacteriol. 2003. PMID: 12896996 Free PMC article.
-
Inactivation of the Pseudomonas putida cytochrome o ubiquinol oxidase leads to a significant change in the transcriptome and to increased expression of the CIO and cbb3-1 terminal oxidases.Environ Microbiol. 2006 Oct;8(10):1764-74. doi: 10.1111/j.1462-2920.2006.01061.x. Environ Microbiol. 2006. PMID: 16958757
-
Transcriptional control of the Pseudomonas putida TOL plasmid catabolic pathways.Mol Microbiol. 1993 Sep;9(5):923-9. doi: 10.1111/j.1365-2958.1993.tb01222.x. Mol Microbiol. 1993. PMID: 7934920 Review.
-
Transcriptional activation of the catechol and chlorocatechol operons: variations on a theme.Gene. 1998 Nov 26;223(1-2):257-67. doi: 10.1016/s0378-1119(98)00366-7. Gene. 1998. PMID: 9858745 Review.
Cited by
-
In-depth characterization of denitrifier communities across different soil ecosystems in the tundra.Environ Microbiome. 2022 Jun 11;17(1):30. doi: 10.1186/s40793-022-00424-2. Environ Microbiome. 2022. PMID: 35690846 Free PMC article.
-
Variability in subpopulation formation propagates into biocatalytic variability of engineered Pseudomonas putida strains.Front Microbiol. 2015 Oct 1;6:1042. doi: 10.3389/fmicb.2015.01042. eCollection 2015. Front Microbiol. 2015. PMID: 26483771 Free PMC article.
-
The Interaction between Plants and Bacteria in the Remediation of Petroleum Hydrocarbons: An Environmental Perspective.Front Microbiol. 2016 Nov 21;7:1836. doi: 10.3389/fmicb.2016.01836. eCollection 2016. Front Microbiol. 2016. PMID: 27917161 Free PMC article. Review.
-
Functional genomics of stress response in Pseudomonas putida KT2440.J Bacteriol. 2006 Jun;188(11):4079-92. doi: 10.1128/JB.00101-06. J Bacteriol. 2006. PMID: 16707699 Free PMC article.
-
Involvement of cyclopropane fatty acids in the response of Pseudomonas putida KT2440 to freeze-drying.Appl Environ Microbiol. 2006 Jan;72(1):472-7. doi: 10.1128/AEM.72.1.472-477.2006. Appl Environ Microbiol. 2006. PMID: 16391080 Free PMC article.
References
-
- Anraku, Y., and R. B. Gennis. 1987. The aerobic respiratory chain of Escherichia coli. Trends Biochem. Sci. 12:262-266.
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl. 1989. Current protocols in molecular biology. John Wiley & Sons, Inc., New York, N.Y.
-
- Bauchop, T., and S. R. Eldsen. 1960. The growth of microorganisms in relation to their energy supply. J. Gen. Microbiol. 23:457-569. - PubMed
-
- Boyer, H. W., and D. Roulland-Dussoix. 1969. A complementation analysis of the restriction and modification of DNA in Escherichia coli. J. Mol. Biol. 41:459-472. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
