

Cell cycle-dependent localization of macroH2A in chromatin of the inactive X chromosome
- PMID: 12082075
- PMCID: PMC2173542
- DOI: 10.1083/jcb.200112074
Cell cycle-dependent localization of macroH2A in chromatin of the inactive X chromosome
Abstract
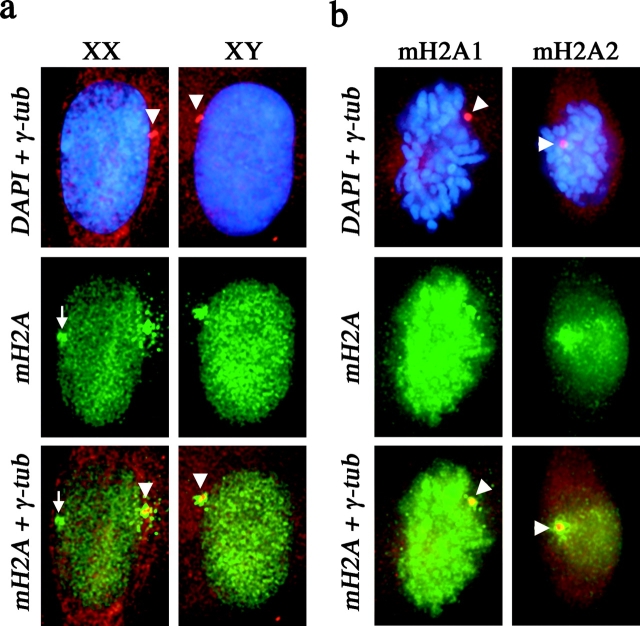
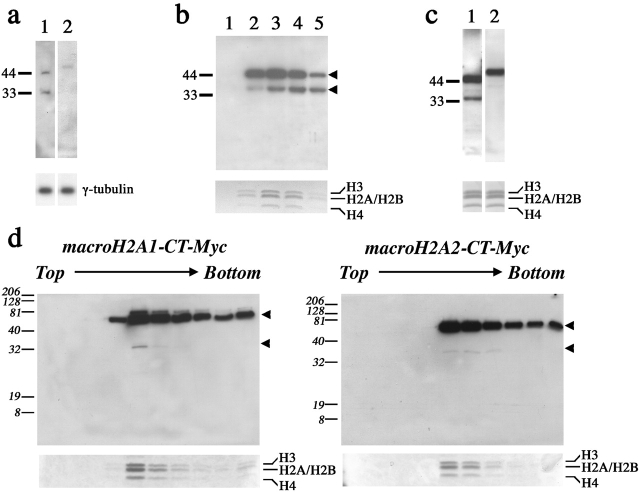
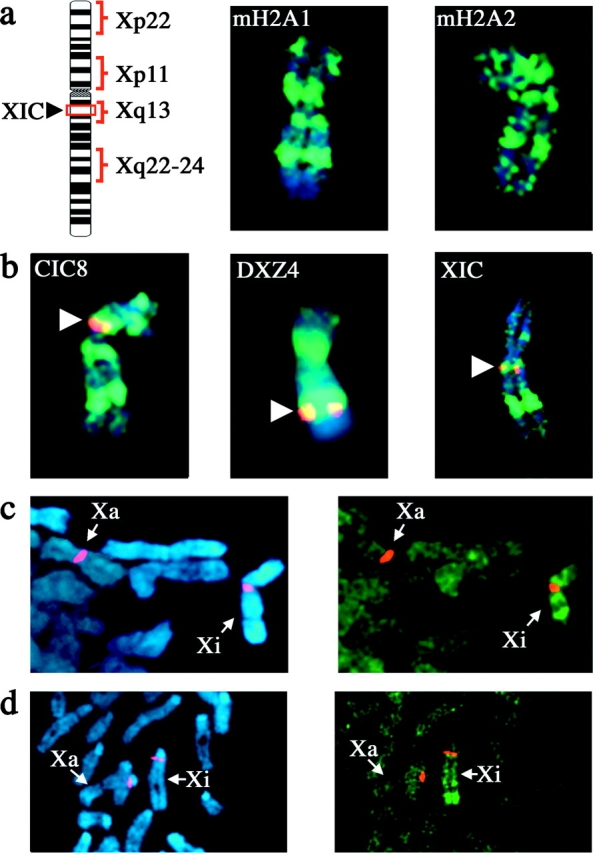
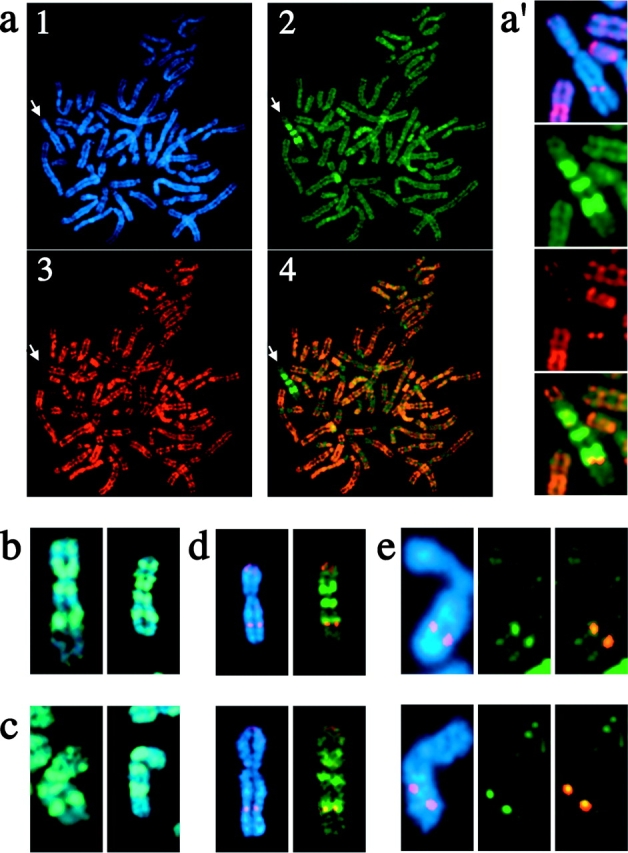
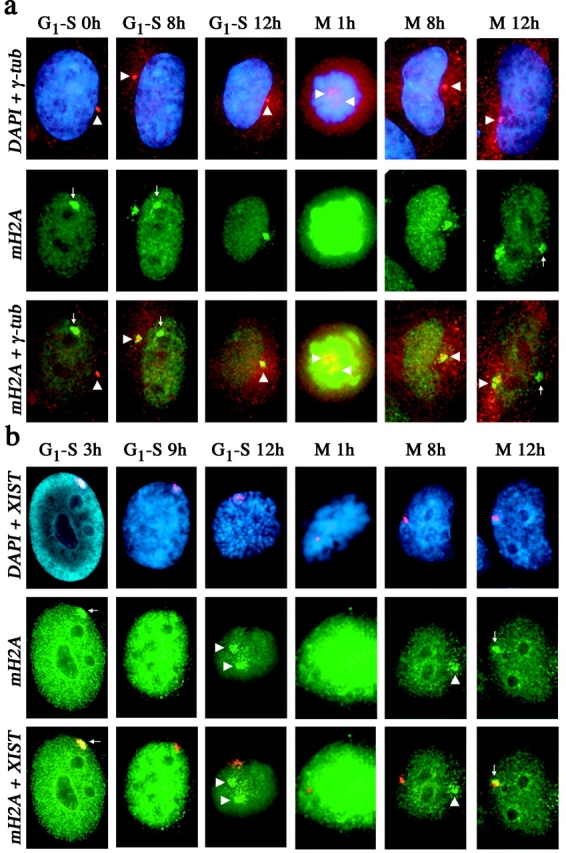
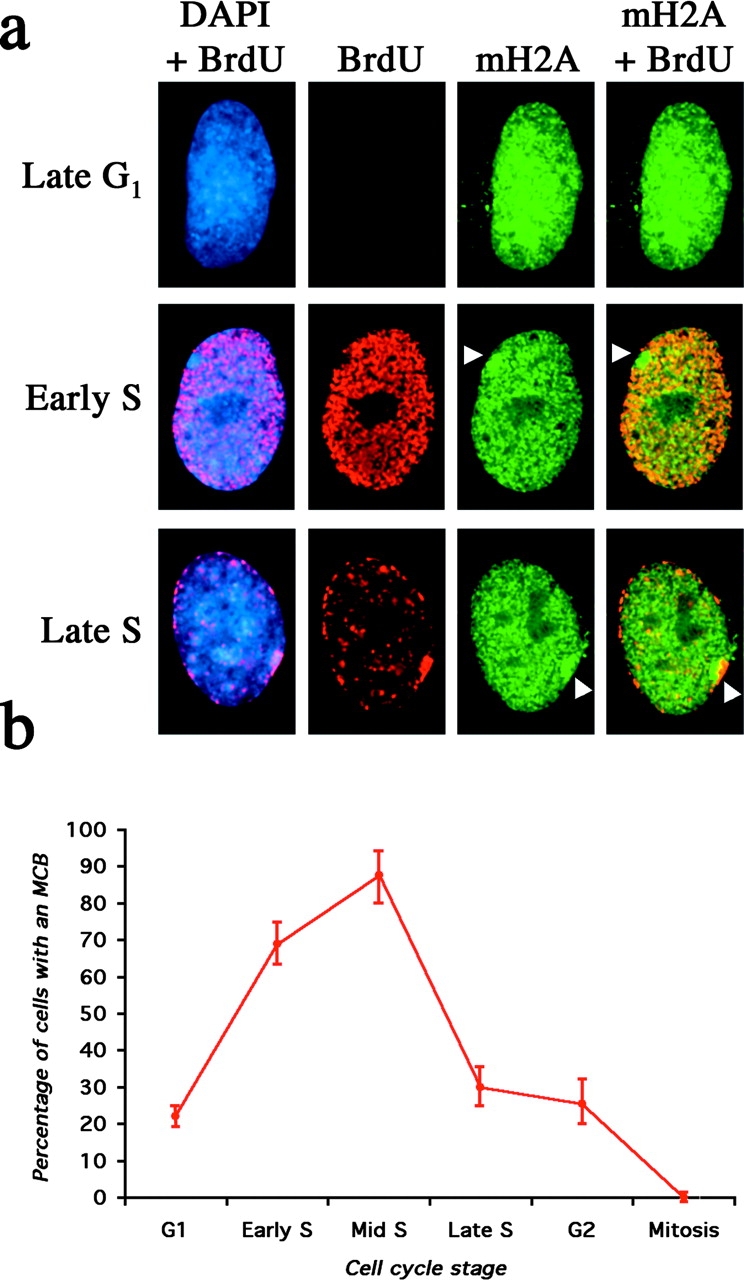
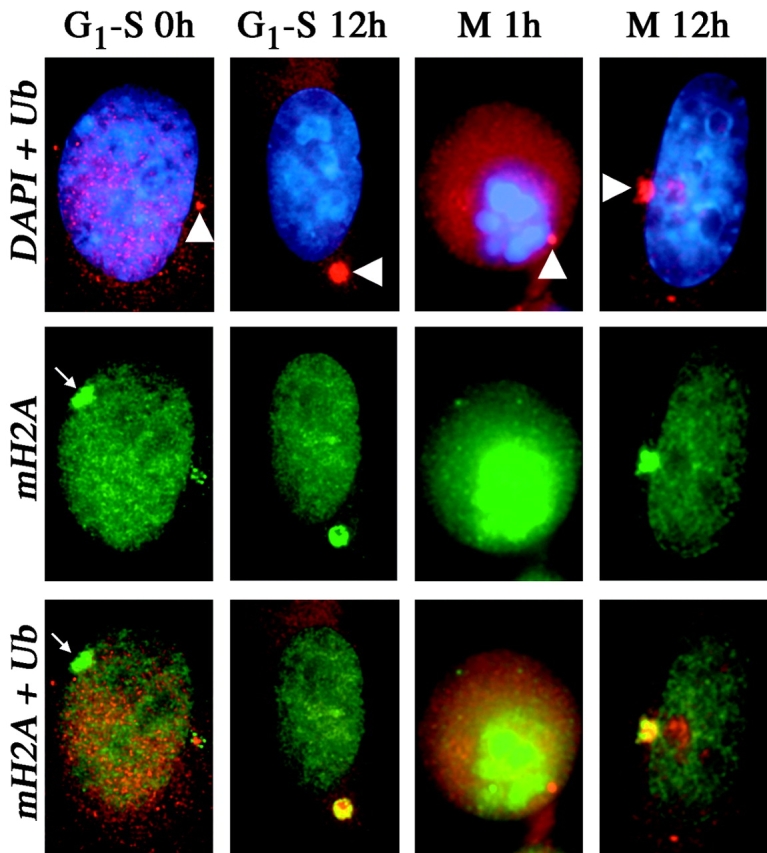
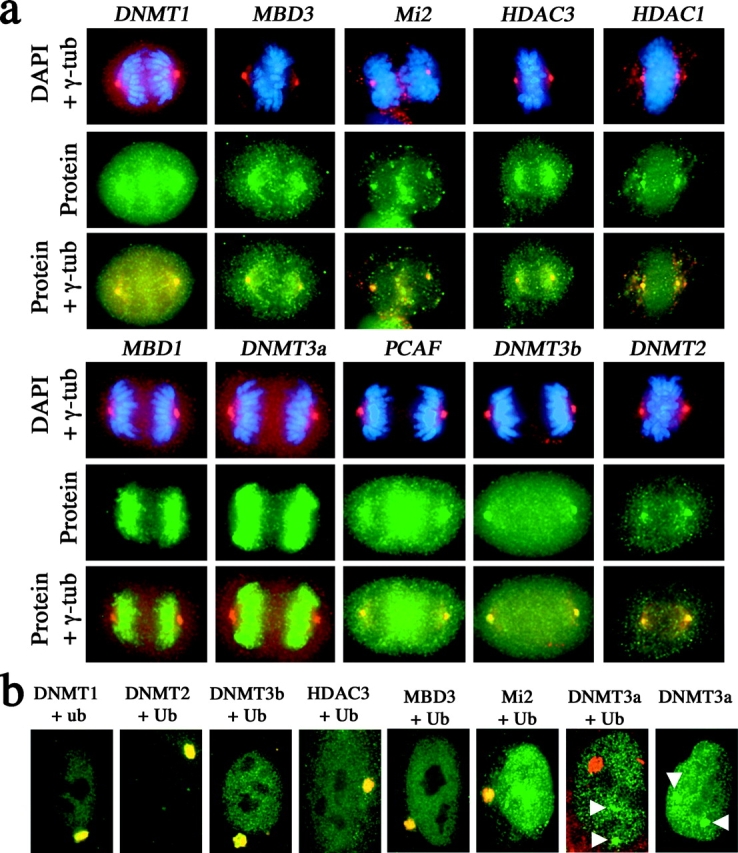
One of several features acquired by chromatin of the inactive X chromosome (Xi) is enrichment for the core histone H2A variant macroH2A within a distinct nuclear structure referred to as a macrochromatin body (MCB). In addition to localizing to the MCB, macroH2A accumulates at a perinuclear structure centered at the centrosome. To better understand the association of macroH2A1 with the centrosome and the formation of an MCB, we investigated the distribution of macroH2A1 throughout the somatic cell cycle. Unlike Xi-specific RNA, which associates with the Xi throughout interphase, the appearance of an MCB is predominantly a feature of S phase. Although the MCB dissipates during late S phase and G2 before reforming in late G1, macroH2A1 remains associated during mitosis with specific regions of the Xi, including at the X inactivation center. This association yields a distinct macroH2A banding pattern that overlaps with the site of histone H3 lysine-4 methylation centered at the DXZ4 locus in Xq24. The centrosomal pool of macroH2A1 accumulates in the presence of an inhibitor of the 20S proteasome. Therefore, targeting of macroH2A1 to the centrosome is likely part of a degradation pathway, a mechanism common to a variety of other chromatin proteins.
Figures
References
-
- Avner, P., and E. Heard. 2001. X-chromosome inactivation: counting, choice and initiation. Nat. Rev. Genet. 2:59–67. - PubMed
-
- Barthelmes, H.U., P. Grue, S. Feineis, T. Straub, and F. Boege. 2000. Active DNA topoisomerase IIα is a component of the salt-stable centrosome core. J. Biol. Chem. 275:38823–38830. - PubMed
-
- Belyaev, N., A.M. Keohane, and B.M. Turner. 1996. Differential underacetylation of histones H2A, H3 and H4 on the inactive X chromosome in human female cells. Hum. Genet. 97:573–578. - PubMed
-
- Bochtler, M., L. Ditzel, M. Groll, C. Hartmann, and R. Huber. 1999. The proteasome. Annu. Rev. Biophys. Biomol. Struct. 28:295–317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
