

Stable expression of calpain 3 from a muscle transgene in vivo: immature muscle in transgenic mice suggests a role for calpain 3 in muscle maturation
- PMID: 12084932
- PMCID: PMC124391
- DOI: 10.1073/pnas.132269299
Stable expression of calpain 3 from a muscle transgene in vivo: immature muscle in transgenic mice suggests a role for calpain 3 in muscle maturation
Abstract
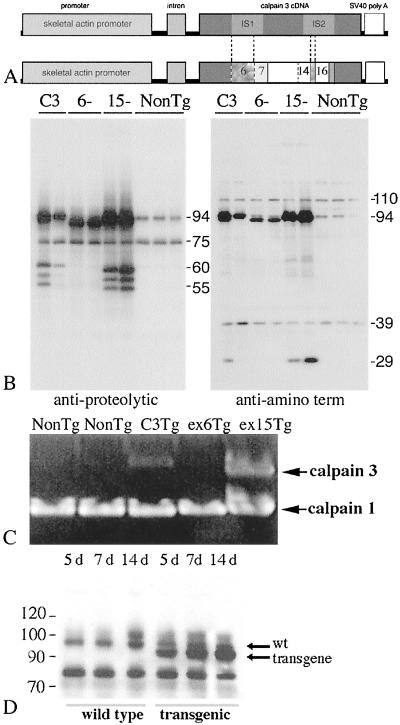
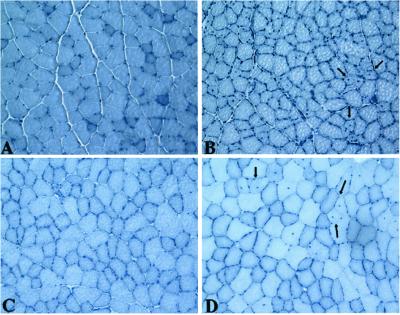
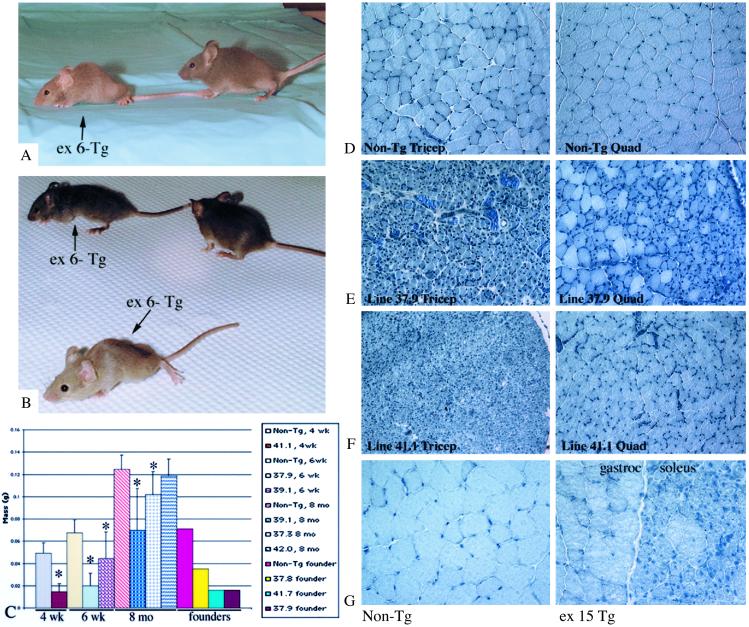
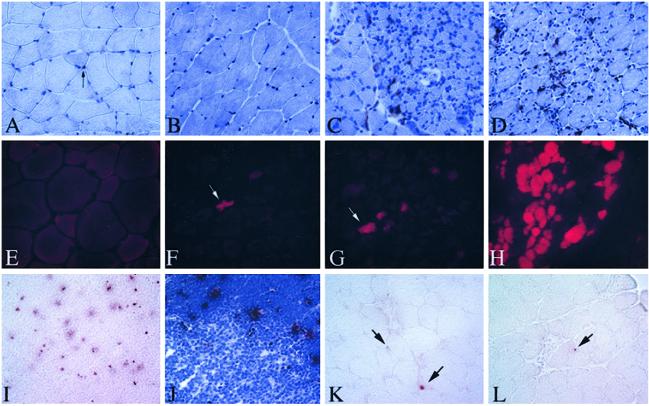
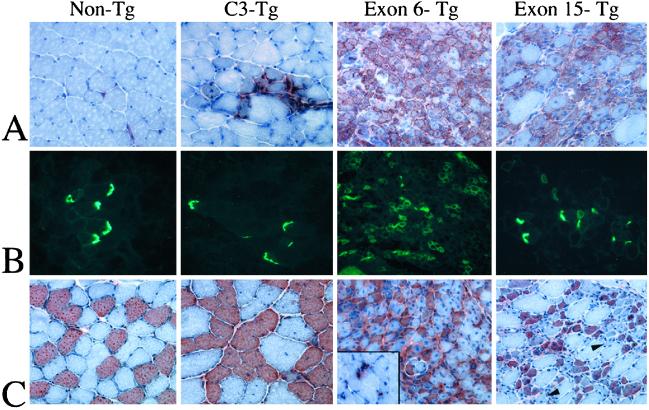
Limb-girdle muscular dystrophy, type 2A (LGMD 2A), is an autosomal recessive disorder that causes late-onset muscle-wasting, and is due to mutations in the muscle-specific protease calpain 3 (C3). Although LGMD 2A would be a feasible candidate for gene therapy, the reported instability of C3 in vitro raised questions about the potential of obtaining a stable, high-level expression of C3 from a transgene in vivo. We have generated transgenic (Tg) mice with muscle-specific overexpression of full-length C3 or C3 isoforms, which arise from alternative splicing, to test whether stable expression of C3 transgenes could occur in vivo. Unexpectedly, we found that full-length C3 can be overexpressed at high levels in vivo, without toxicity. In addition, we found that Tg expressing C3 lacking exon 6, an isoform expressed embryonically, have muscles that resemble regenerating or developing muscle. Tg expressing C3 lacking exon 15 shared this morphology in the soleus, but not other muscles. Assays of inflammation or muscle membrane damage indicated that the Tg muscles were not degenerative, suggesting that the immature muscle resulted from a developmental block rather than degeneration and regeneration. These studies show that C3 can be expressed stably in vivo from a transgene, and indicate that alternatively spliced C3 isoforms should not be used in gene-therapy applications because they impair proper muscle development.
Figures
References
-
- Richard I, Broux O, Allamand V, Fougerousse F, Chiannilkulchai N, Bourg N, Brenguier L, Devaud C, Pasturaud P, Roudaut C, et al. Cell. 1995;81:27–40. - PubMed
-
- Sorimachi H, Suzuki K. J Biochem. 2001;129:653–664. - PubMed
-
- Sorimachi H, Toyama-Sorimachi N, Saido T C, Kawasaki H, Sugita H, Miyasaka M, Arahata K, Ishiura S, Suzuki K. J Biol Chem. 1993;268:10593–10605. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
