

The mitochondrial DNA polymerase as a target of oxidative damage
- PMID: 12087165
- PMCID: PMC117047
- DOI: 10.1093/nar/gkf392
The mitochondrial DNA polymerase as a target of oxidative damage
Abstract
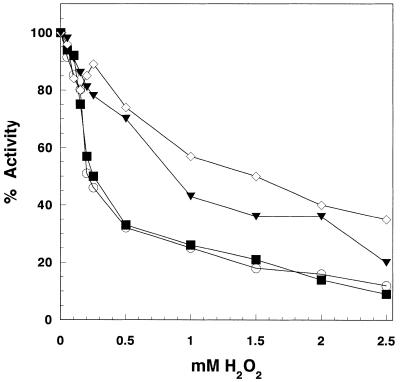
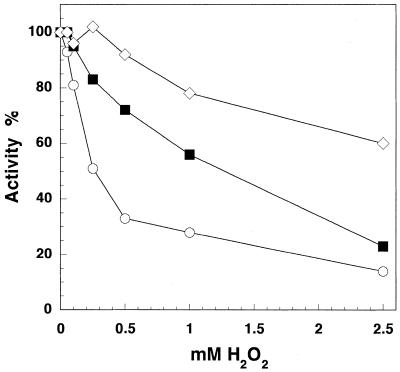
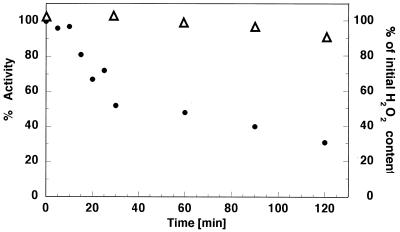
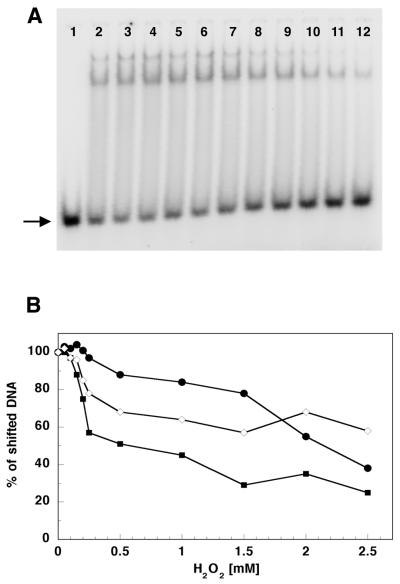
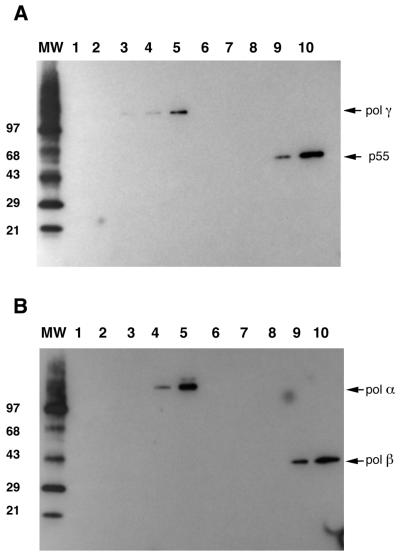
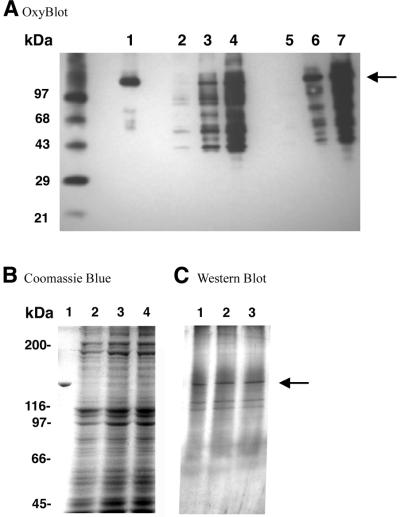
The mitochondrial respiratory chain is a source of reactive oxygen species (ROS) that are responsible for oxidative modification of biomolecules, including proteins. Due to its association with mitochondrial DNA, DNA polymerase gamma (pol gamma) is in an environment to be oxidized by hydrogen peroxide and hydroxyl radicals that may be generated in the presence of iron ions associated with DNA. We tested whether human pol gamma was a possible target of ROS with H2O2 and investigated the effect on the polymerase activities and DNA binding efficiency. A 1 h treatment with 250 microM H2O2 significantly inhibited DNA polymerase activity of the p140 subunit and lowered its DNA binding efficiency. Addition of p55 to the p140 catalytic subunit prior to H2O2 treatment offered protection from oxidative inactivation. Oxidatively modified amino acid residues in pol gamma resulting from H2O2 treatment were observed in vitro as well as in vivo, in SV40-transfected human fibroblasts. Pol gamma was detected as one of the major oxidized mitochondrial matrix proteins, with a detectable decline in polymerase activity. These results suggest pol gamma as a target of oxidative damage, which may result in a reduction in mitochondrial DNA replication and repair capacities.
Figures
References
-
- Michikawa Y., Mazzucchelli,F., Bresolin,N., Scarlato,G. and Attardi,G. (1999) Aging-dependent large accumulation of point mutations in the human mtDNA control region for replication. Science, 286, 774–779. - PubMed
-
- Chance B., Sies,H. and Boveris,A. (1979) Hydroperoxide metabolism in mammalian organs. Physiol. Rev., 59, 527–605. - PubMed
-
- Ballinger S.W., Van Houten,B., Jin,G.F., Conklin,C.A. and Godley,B.F. (1999) Hydrogen peroxide causes significant mitochondrial DNA damage in human RPE cells. Exp. Eye Res., 68, 765–772. - PubMed
-
- Salazar J.J. and Van Houten,B. (1997) Preferential mitochondrial DNA injury caused by glucose oxidase as a steady generator of hydrogen peroxide in human fibroblasts. Mutat. Res., 385, 139–149. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
