

Premature termination of RNA polymerase II mediated transcription of a seed protein gene in Schizosaccharomyces pombe
- PMID: 12087180
- PMCID: PMC117036
- DOI: 10.1093/nar/gkf334
Premature termination of RNA polymerase II mediated transcription of a seed protein gene in Schizosaccharomyces pombe
Abstract
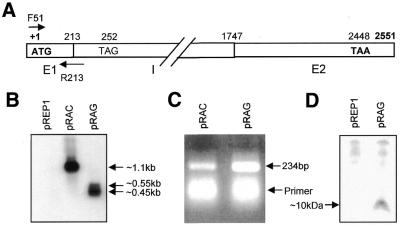
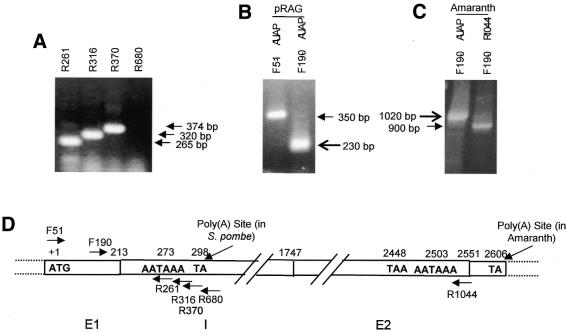
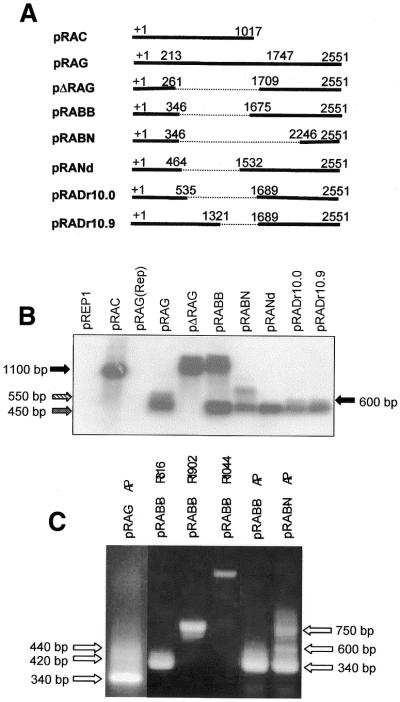
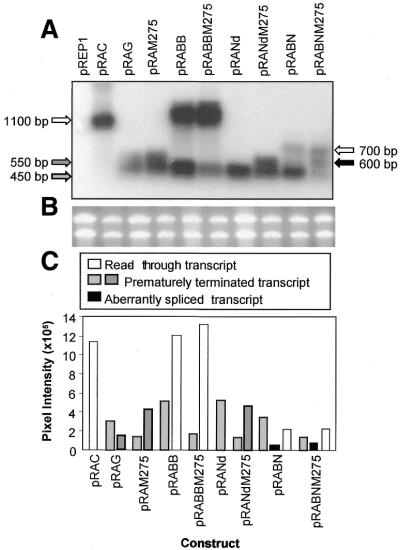
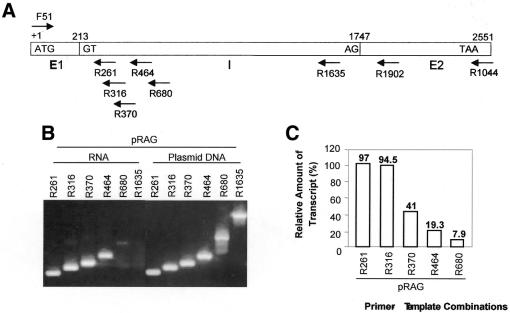
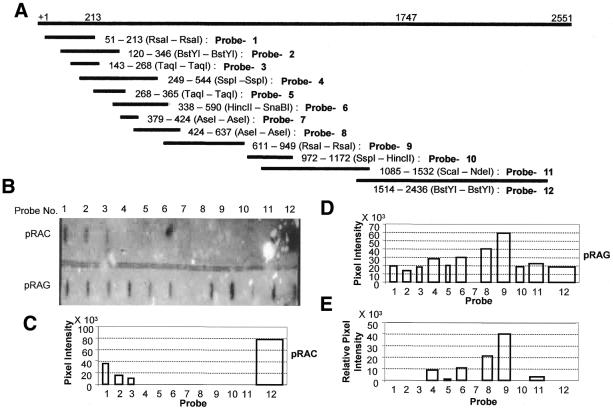
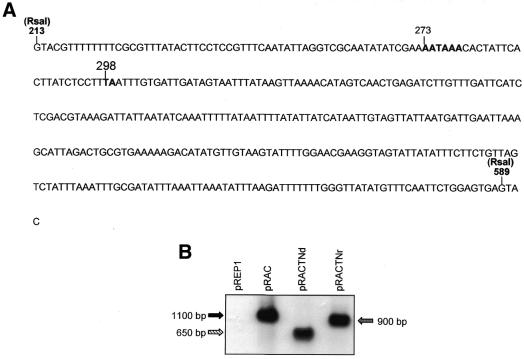
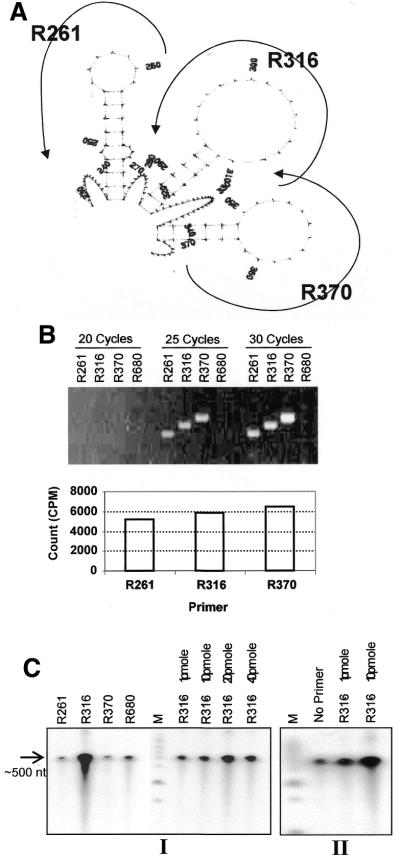
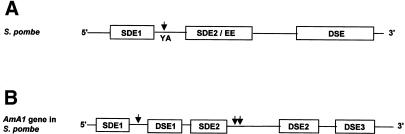
The poly(A) signal and downstream elements with transcriptional pausing activity play an important role in termination of RNA polymerase II transcription. We show that an intronic sequence derived from the plant seed protein gene (AmA1) specifically acts as a transcriptional terminator in the fission yeast, Schizosaccharomyces pombe. The 3'-end points of mRNA encoded by the AmA1 gene were mapped at different positions in S.pombe and in native cells of Amaranthus hypochondriacus. Deletion analyses of the AmA1 intronic sequence revealed that multiple elements essential for proper transcriptional termination in S.pombe include two site-determining elements (SDEs) and three downstream sequence elements. RT-PCR analyses detected transcripts up to the second SDE. This is the first report showing that the highly conserved mammalian poly(A) signal, AAUAAA, is also functional in S.pombe. The poly(A) site was determined as Y(A) both in native and heterologous systems but at different positions. Deletion of these cis-elements abolished 3'-end processing in S.pombe and a single point mutation in this motif reduced the activity by 70% while enhancing activity at downstream SDE. These results indicate that the bipartite sequence elements as signals for 3'-end processing in fission yeast act in tandem with other cis-acting elements. A comparison of these elements in the AmA1 intron that function as a transcriptional terminator in fission yeast with that of its native genes showed that both require an AT-rich distal and proximal upstream element. However, these sequences are not identical. Transcription run-on analysis indicates that elongating RNA polymerase II molecules accumulate over these pause signals, maximal at 611-949 nt. Furthermore, we demonstrate that the AmA1 intronic terminator sequence acts in a position-independent manner when placed within another gene.
Figures
Similar articles
-
Transcriptional termination signals for RNA polymerase II in fission yeast.EMBO J. 1997 Jun 16;16(12):3633-43. doi: 10.1093/emboj/16.12.3633. EMBO J. 1997. PMID: 9218804 Free PMC article.
-
RNA 3' end signals of the S.pombe ura4 gene comprise a site determining and efficiency element.EMBO J. 1994 May 15;13(10):2441-51. doi: 10.1002/j.1460-2075.1994.tb06529.x. EMBO J. 1994. PMID: 8194534 Free PMC article.
-
Plant pre-mRNA splicing in fission yeast, Schizosaccharomyces pombe.Biochem Biophys Res Commun. 2002 May 17;293(4):1209-16. doi: 10.1016/S0006-291X(02)00364-9. Biochem Biophys Res Commun. 2002. PMID: 12054504
-
Comparison of the RNA polymerase III transcription machinery in Schizosaccharomyces pombe, Saccharomyces cerevisiae and human.Nucleic Acids Res. 2001 Jul 1;29(13):2675-90. doi: 10.1093/nar/29.13.2675. Nucleic Acids Res. 2001. PMID: 11433012 Free PMC article. Review.
-
Transcription termination and 3' processing: the end is in site!Cell. 1985 Jun;41(2):349-59. doi: 10.1016/s0092-8674(85)80007-6. Cell. 1985. PMID: 2580642 Review. No abstract available.
Cited by
-
A complex gene regulatory mechanism that operates at the nexus of multiple RNA processing decisions.Nat Struct Mol Biol. 2009 Mar;16(3):255-64. doi: 10.1038/nsmb.1556. Epub 2009 Feb 8. Nat Struct Mol Biol. 2009. PMID: 19198588 Free PMC article.
-
Effect of codon optimization on expression levels of a functionally folded malaria vaccine candidate in prokaryotic and eukaryotic expression systems.Infect Immun. 2003 Sep;71(9):4961-9. doi: 10.1128/IAI.71.9.4961-4969.2003. Infect Immun. 2003. PMID: 12933838 Free PMC article.
-
Next-generation protein-rich potato expressing the seed protein gene AmA1 is a result of proteome rebalancing in transgenic tuber.Proc Natl Acad Sci U S A. 2010 Oct 12;107(41):17533-8. doi: 10.1073/pnas.1006265107. Epub 2010 Sep 20. Proc Natl Acad Sci U S A. 2010. PMID: 20855595 Free PMC article.
References
-
- Birse C.E., Minvielle-Sebastia,L., Lee,B.A., Keller,W. and Proudfoot,N.J. (1998) Coupling termination of transcription to messenger RNA maturation in yeast. Science, 280, 298–301. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
