

Continuous porcine cell lines developed from alveolar macrophages: partial characterization and virus susceptibility
- PMID: 12088830
- PMCID: PMC7119708
- DOI: 10.1016/s0166-0934(02)00085-x
Continuous porcine cell lines developed from alveolar macrophages: partial characterization and virus susceptibility
Abstract
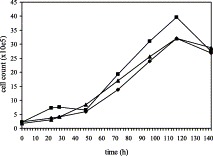
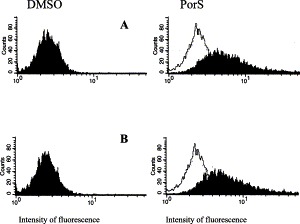
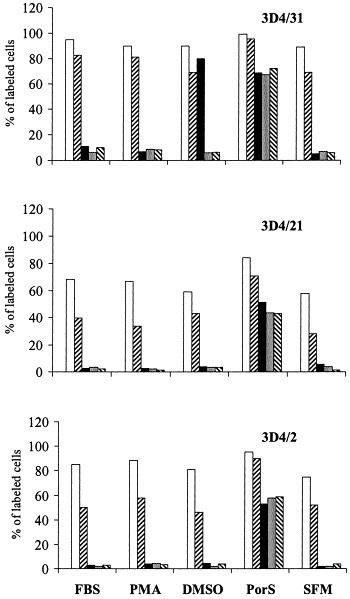
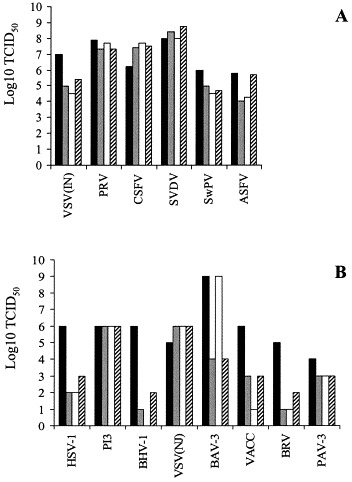
Porcine monomyeloid cell lines were established following transfection of primary porcine alveolar macrophage cultures with plasmid pSV3neo, carrying genes for neomycin resistance and SV40 large T antigen. The parental clone 3D4 exhibited a relatively rapid doubling time (25.5 h), high plating efficiency and mixed phenotype with respect to growth on a solid support. Single cell cloning of the 3D4 parent resulted in establishment of several cell lines; three of them designated 3D4/2, 3D4/21 and 3D4/31 were selected for further characterization. All three clones supported the replication of vesicular stomatitis virus (VSV), pseudorabies virus (PRV), classical swine fever virus (CSFV), swine vesicular disease virus (SVDV), swine poxvirus, African swine fever virus (ASFV), herpes simplex virus (HSV), parainfluenza virus, bovine adenovirus (BAV), vaccinia virus (VV), and porcine adenovirus (PAV). Under the conditions tested the cells did not support replication of porcine reproductive and respiratory syndrome virus (PRRSV). The swine myeloid character was confirmed for all three clones by fluorescence activated cell scanning (FACS) analysis using monoclonal antibodies 74-22-15 and DH59B, which recognize the pan-myeloid antigen cluster SWC3a. A subpopulation of each cell line was positive for nonspecific esterase activity and phagocytic activity to varying degrees depending on the media formulation. Cells from all three lines exhibited anchorage dependent growth when maintained in RPMI 1640 medium supplemented with 5-15% fetal bovine serum (FBS) and nonessential amino acids. Propagation in commercially formulated serum free media resulted in colony formation and growth in suspension. The addition of dimethyl sulfoxide (DMSO) or phorbol 12-myristate 13-acetate (PMA) to serum free media restored cell attachment. DMSO was also able to induce expression of CD14 monocyte marker in the 3D4/31 cell line maintained in FBS containing medium, as determined by FACS with monoclonal antibody CAM36A. Supplementation of RPMI medium with 10% porcine serum upregulated the expression of CD14 and induced expression of porcine macrophage markers recognized by antibodies 2B10 and 2G6 (Vet. Immunol. Immunopathol. 74 (2000) 163) in all three cell lines. The porcine myelomonocytic cell lines obtained may have a wide variety of applications in porcine virology and immunology.
Figures
Similar articles
-
A simple and efficient method for the generation of a porcine alveolar macrophage cell line for high-efficiency Porcine reproductive and respiratory syndrome virus 2 infection.J Virol Methods. 2019 Dec;274:113727. doi: 10.1016/j.jviromet.2019.113727. Epub 2019 Sep 4. J Virol Methods. 2019. PMID: 31493424
-
Establishment of transformed swine fibroblast cell lines using SV40 large T antigen.Arch Virol. 1990;115(3-4):227-37. doi: 10.1007/BF01310532. Arch Virol. 1990. PMID: 2175590
-
Generation of a porcine alveolar macrophage cell line for the growth of porcine reproductive and respiratory syndrome virus.J Virol Methods. 2010 Feb;163(2):410-5. doi: 10.1016/j.jviromet.2009.11.003. Epub 2009 Nov 10. J Virol Methods. 2010. PMID: 19900480
-
The role of pulmonary intravascular macrophages in porcine reproductive and respiratory syndrome virus infection.Anim Health Res Rev. 2000 Dec;1(2):95-102. doi: 10.1017/s1466252300000086. Anim Health Res Rev. 2000. PMID: 11708601 Review.
-
Porcine Macrophage Markers and Populations: An Update.Cells. 2023 Aug 19;12(16):2103. doi: 10.3390/cells12162103. Cells. 2023. PMID: 37626913 Free PMC article. Review.
Cited by
-
An immortalized porcine macrophage cell line competent for the isolation of African swine fever virus.Sci Rep. 2021 Feb 26;11(1):4759. doi: 10.1038/s41598-021-84237-2. Sci Rep. 2021. PMID: 33637799 Free PMC article.
-
Development and characterization of two porcine monocyte-derived macrophage cell lines.Results Immunol. 2013;3:26-32. doi: 10.1016/j.rinim.2013.03.001. Results Immunol. 2013. PMID: 23610747 Free PMC article.
-
A Multi-Laboratory Comparison of Methods for Detection and Quantification of African Swine Fever Virus.Pathogens. 2022 Mar 7;11(3):325. doi: 10.3390/pathogens11030325. Pathogens. 2022. PMID: 35335649 Free PMC article.
-
Breed Differences in PCV2 Uptake and Disintegration in Porcine Monocytes.Viruses. 2018 Oct 15;10(10):562. doi: 10.3390/v10100562. Viruses. 2018. PMID: 30326643 Free PMC article.
-
Porcine Reproductive and Respiratory Syndrome Virus Nonstructural Protein 1 Beta Interacts with Nucleoporin 62 To Promote Viral Replication and Immune Evasion.J Virol. 2019 Jun 28;93(14):e00469-19. doi: 10.1128/JVI.00469-19. Print 2019 Jul 15. J Virol. 2019. PMID: 31043527 Free PMC article.
References
-
- Alvarez B., Sanchez C., Bullido R., Marina A., Lunney J., Alonso F., Ezquerra A., Dominguez J. A porcine cell surface receptor identified by monoclonal antibodies to SWC3 is a member of the signal regulatory protein family and associates with protein-tyrosine phosphatase SHP-1. Tissue Antigens. 2000;55:342–351. - PubMed
-
- Basta S., Knoetig S.M., Spagnuolo-Weaver M., Allan G., McCullough K.C. Modulation of monocytic cell activity and virus susceptibility during differentiation into macrophages. J. Immunol. 1999;162:3961–3969. - PubMed
-
- Benfield, D.A., Collins, J.E., Dee, S.A., Halbur, P.G., Joo, H.S., Lager, K.M., Mengeling, W.L., Murtaugh, M.P., Rossow, K.D., Stevenson, G.W., Zimmerman, J.J., 1999. Porcine reproductive and respiratory syndrome. In: Straw, B.E., D'Allaire, S., Mengeling, W.L., Taylor, D.J. (Eds.) Diseases of Swine, 8th edition, Iowa State University Press, Ames, Iowa, pp. 201–232.
-
- Berndt A., Heller M., Methner U., Kosmel H., Muller G. Monoclonal antibodies against porcine macrophages. Vet. Immunol. Immunopathol. 2000;74:163–177. - PubMed
-
- Brodersen R., Bijlsma F., Gori K., Jensen K.T., Chen W., Dominguez J., Haverson K., Moore P.F., Saalmuller A., Sachs D., Slierendrecht W.J., Stokes C., Vainio O., Zuckermann F., Aasted B. Analysis of the immunological cross reactivities of 213 well characterized monoclonal antibodies with specificities against various leukocyte surface antigens of human and 11 animal species. Vet. Immunol. Immunopathol. 1998;64:1–13. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
