

Improving the pathogenicity of a nematode-trapping fungus by genetic engineering of a subtilisin with nematotoxic activity
- PMID: 12089022
- PMCID: PMC126817
- DOI: 10.1128/AEM.68.7.3408-3415.2002
Improving the pathogenicity of a nematode-trapping fungus by genetic engineering of a subtilisin with nematotoxic activity
Abstract
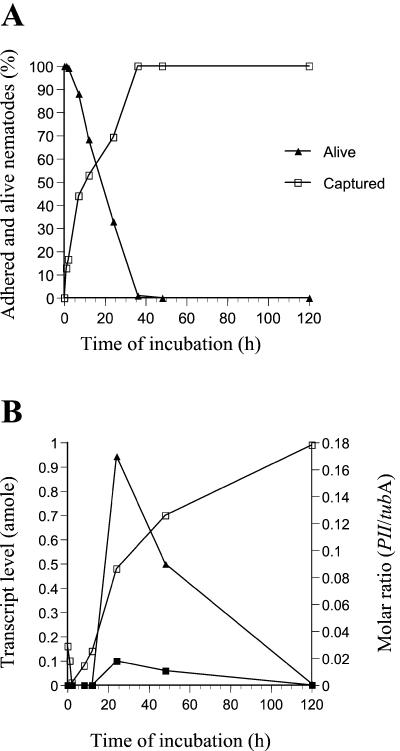
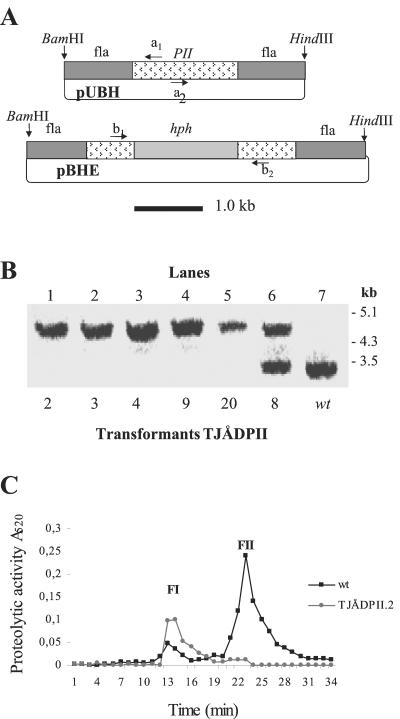
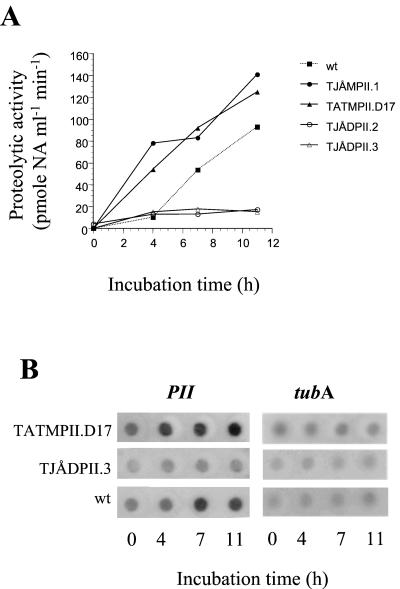
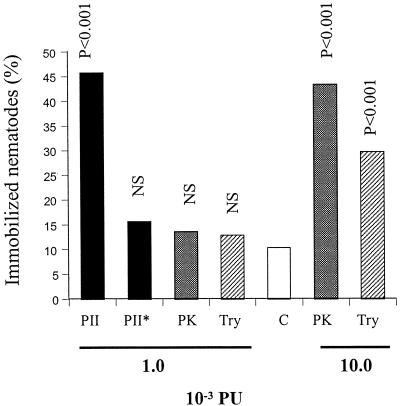
Nematophagous fungi are soil-living fungi that are used as biological control agents of plant and animal parasitic nematodes. Their potential could be improved by genetic engineering, but the lack of information about the molecular background of the infection has precluded this development. In this paper we report that a subtilisin-like extracellular serine protease designated PII is an important pathogenicity factor in the common nematode-trapping fungus Arthrobotrys oligospora. The transcript of PII was not detected during the early stages of infection (adhesion and penetration), but high levels were expressed concurrent with the killing and colonization of the nematode. Disruption of the PII gene by homologous recombination had a limited effect on the pathogenicity of the fungus. However, mutants containing additional copies of the PII gene developed a higher number of infection structures and had an increased speed of capturing and killing nematodes compared to the wild type. The paralyzing activity of PII was verified by demonstrating that a heterologous-produced PII (in Aspergillus niger) had a nematotoxic activity when added to free-living nematodes. The toxic activity of PII was significantly higher than that of other commercially available serine proteases. This is the first report showing that genetic engineering can be used to improve the pathogenicity of a nematode-trapping fungus. In the future it should be possible to express recombinant subtilisins with nematicidal activity in other organisms that are present in the habitat of parasitic nematodes (e.g., host plant).
Figures
References
-
- Åhman, J., B. Ek, L. Rask, and A. Tunlid. 1996. Sequence analysis and regulation of a gene encoding a cuticle-degrading serine protease from the nematophagous fungus Arthrobotrys oligospora. Microbiology 142:1605-1616. - PubMed
-
- Bennet, J. W., and L. L. Lasure. 1991. Growth media, p. 444-445. In J. W. Bennet and L. L. Lasure (ed.), More gene manipulations in fungi. Academic Press, San Diego, Calif.
-
- Bonants, P. J. M., P. F. L. Fitters, H. Thijs, E. Den-Belder, C. Waalwijk, and J. W. D. M. Henfling. 1995. A basic serine protease from Paecilomyces lilacinus with biological activity against Meloidogyne hapla eggs. Microbiology 141:775-784. - PubMed
-
- Carrol, A. M., J. A. Sweigard, and B. Valent. 1995. Improved vectors for selecting resistance to hygromycin. Fungal Gen. News 41:22.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
