

Rapid detection and quantification of RNA of Ebola and Marburg viruses, Lassa virus, Crimean-Congo hemorrhagic fever virus, Rift Valley fever virus, dengue virus, and yellow fever virus by real-time reverse transcription-PCR
- PMID: 12089242
- PMCID: PMC120575
- DOI: 10.1128/JCM.40.7.2323-2330.2002
Rapid detection and quantification of RNA of Ebola and Marburg viruses, Lassa virus, Crimean-Congo hemorrhagic fever virus, Rift Valley fever virus, dengue virus, and yellow fever virus by real-time reverse transcription-PCR
Abstract
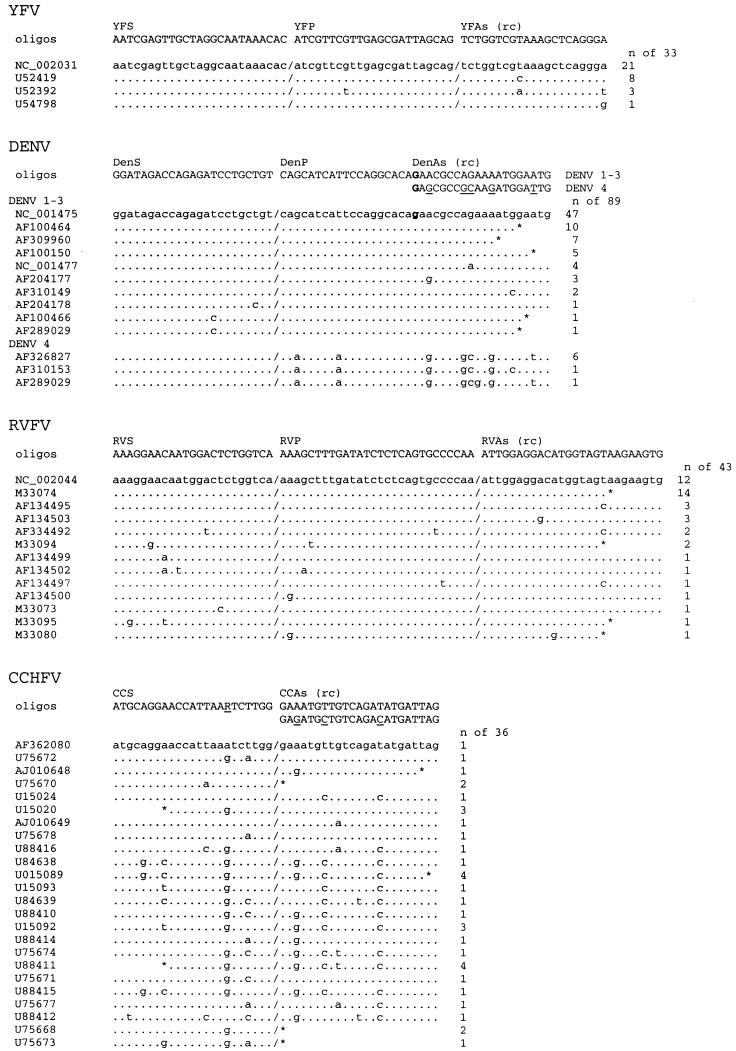
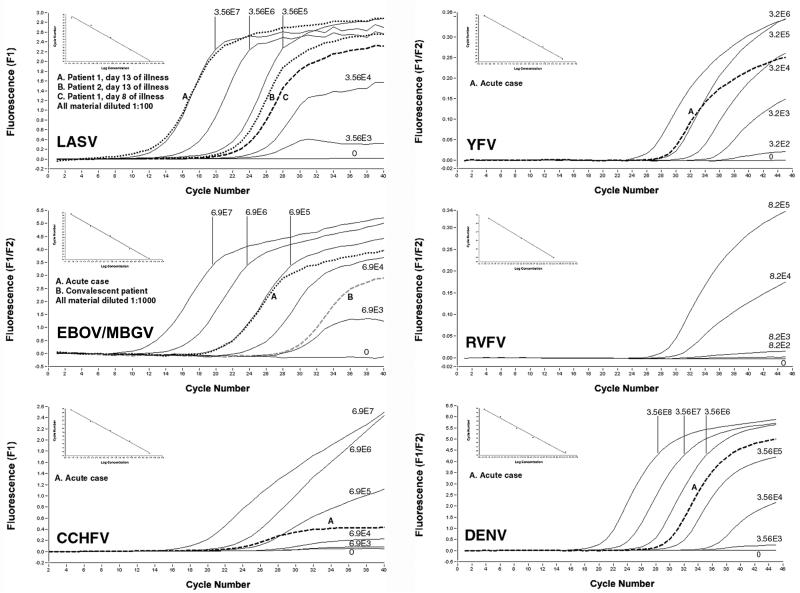
Viral hemorrhagic fevers (VHFs) are acute infections with high case fatality rates. Important VHF agents are Ebola and Marburg viruses (MBGV/EBOV), Lassa virus (LASV), Crimean-Congo hemorrhagic fever virus (CCHFV), Rift Valley fever virus (RVFV), dengue virus (DENV), and yellow fever virus (YFV). VHFs are clinically difficult to diagnose and to distinguish; a rapid and reliable laboratory diagnosis is required in suspected cases. We have established six one-step, real-time reverse transcription-PCR assays for these pathogens based on the Superscript reverse transcriptase-Platinum Taq polymerase enzyme mixture. Novel primers and/or 5'-nuclease detection probes were designed for RVFV, DENV, YFV, and CCHFV by using the latest DNA database entries. PCR products were detected in real time on a LightCycler instrument by using 5'-nuclease technology (RVFV, DENV, and YFV) or SybrGreen dye intercalation (MBGV/EBOV, LASV, and CCHFV). The inhibitory effect of SybrGreen on reverse transcription was overcome by initial immobilization of the dye in the reaction capillaries. Universal cycling conditions for SybrGreen and 5'-nuclease probe detection were established. Thus, up to three assays could be performed in parallel, facilitating rapid testing for several pathogens. All assays were thoroughly optimized and validated in terms of analytical sensitivity by using in vitro-transcribed RNA. The >or=95% detection limits as determined by probit regression analysis ranged from 1,545 to 2,835 viral genome equivalents/ml of serum (8.6 to 16 RNA copies per assay). The suitability of the assays was exemplified by detection and quantification of viral RNA in serum samples of VHF patients.
Figures
References
-
- Anonymous. 2000. Lassa fever imported to England. Commun. Dis. Rep. CDR Wkly. 10:99. - PubMed
-
- Anonymous. 2000. Lassa fever, imported case, Netherlands. Wkly. Epidemiol. Rec. 75:265.. - PubMed
-
- Anonymous. 2001. Outbreak of Ebola haemorrhagic fever, Uganda, August 2000-January 2001. Wkly. Epidemiol. Rec. 76:41-46. - PubMed
-
- Bwaka, M. A., M. J. Bonnet, P. Calain, R. Colebunders, A. De Roo, Y. Guimard, K. R. Katwiki, K. Kibadi, M. A. Kipasa, K. J. Kuvula, B. B. Mapanda, M. Massamba, K. D. Mupapa, J. J. Muyembe-Tamfum, E. Ndaberey, C. J. Peters, P. E. Rollin, and E. Van den Enden. 1999.. Ebola hemorrhagic fever in Kikwit, Democratic Republic of the Congo: clinical observations in 103 patients. J. Infect. Dis. 179(Suppl. 1):S1-S7. - PubMed
-
- Callahan, J. D., S. J. Wu, A. Dion-Schultz, B. E. Mangold, L. F. Peruski, D. M. Watts, K. R. Porter, G. R. Murphy, W. Suharyono, C. C. King, C. G. Hayes, and J. J. Temenak. 2001. Development and evaluation of serotype- and group-specific fluorogenic reverse transcriptase PCR (TaqMan) assays for dengue virus. J. Clin. Microbiol. 39:4119-4124. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
