

GH3, a novel proapoptotic domain in Drosophila Grim, promotes a mitochondrial death pathway
- PMID: 12093734
- PMCID: PMC126101
- DOI: 10.1093/emboj/cdf354
GH3, a novel proapoptotic domain in Drosophila Grim, promotes a mitochondrial death pathway
Abstract
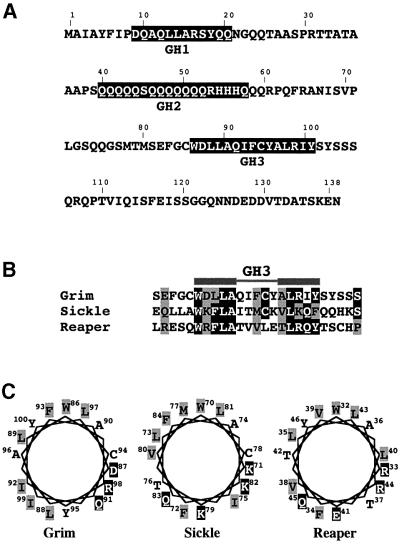
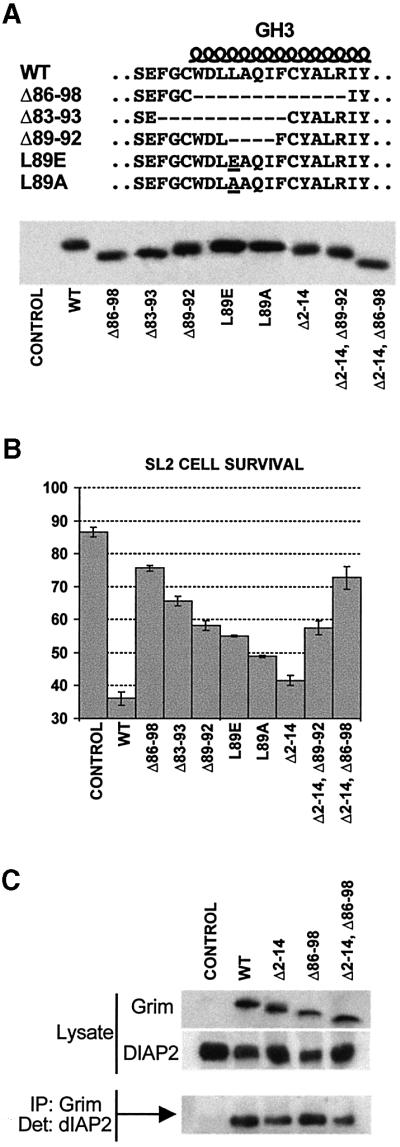
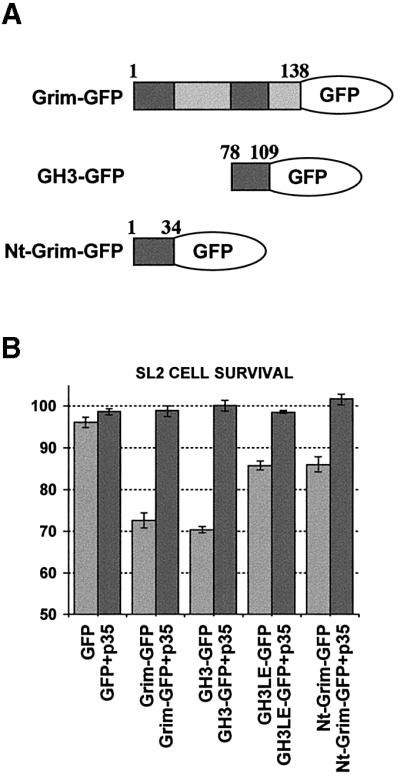
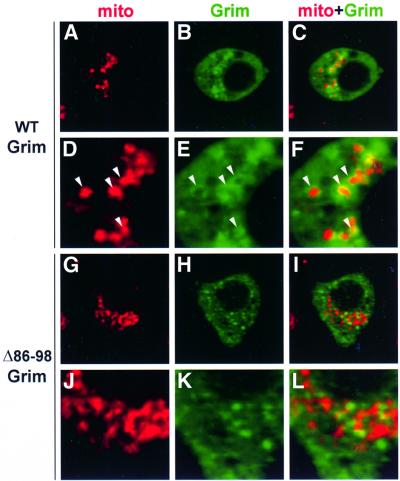
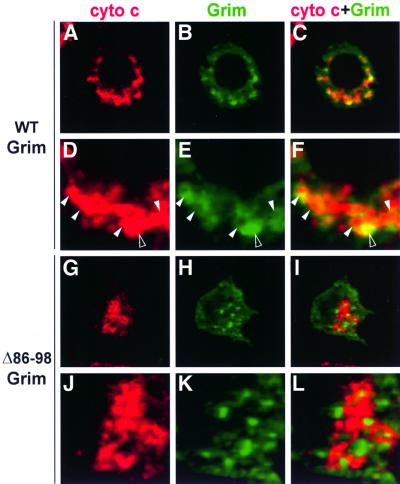
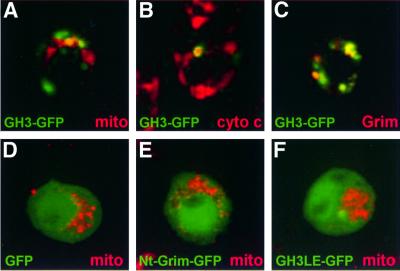
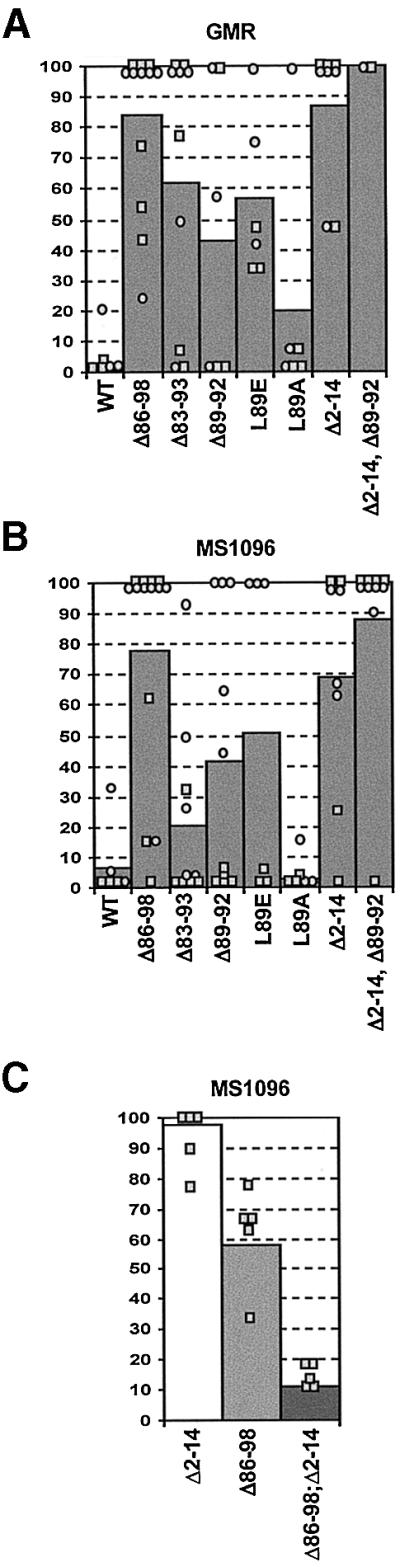
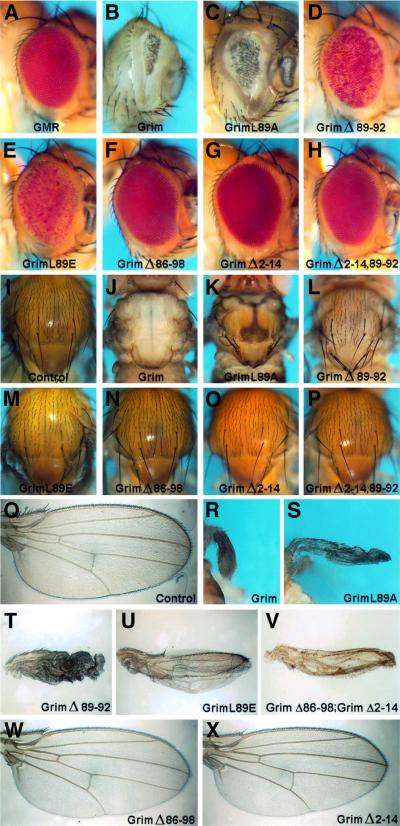
Grim encodes a protein required for programmed cell death in Drosophila. The Grim N-terminus induces apoptosis by disrupting IAP blockage of caspases; however, N-terminally-deleted Grim retains pro apoptotic activity. We describe GH3, a 15 amino acid internal Grim domain absolutely required for its proapoptotic activity and sufficient to induce cell death when fused to heterologous carrier proteins. A GH3 homology region is present in the Drosophila proapoptotic proteins Reaper and Sickle. The GH3 domain and the homologous regions in Reaper and Sickle are predicted to be structured as amphipathic alpha-helixes. During apoptosis induction, Grim colocalizes with mitochondria and cytochrome c in a GH3-dependent but N-terminal- and caspase activity-independent manner. When Grim is overexpressed in vivo, both the N-terminal and the GH3 domains are equally necessary, and cooperate for apoptosis induction. The N-terminal and GH3 Grim domains thus activate independent apoptotic pathways that synergize to induce programmed cell death efficiently.
Figures
References
-
- Abrams J.M. (1999) An emerging blueprint for apoptosis in Drosophila. Trends Cell Biol., 9, 435–440. - PubMed
-
- Aravind L., Dixit,V.M. and Koonin,E.V. (1999) The domains of death: evolution of the apoptosis machinery. Trends Biochem. Sci., 24, 47–53. - PubMed
-
- Ashburner M. (1989) Drosophila: A Laboratory Handbook. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 1017–1063.
-
- Bergmann A., Agapite,J., McCall,K. and Steller,H. (1998) The Drosophila gene hid is a direct molecular target of Ras-dependent survival signaling. Cell, 95, 331–341. - PubMed
-
- Brachmann C.B., Jassim,O.W., Wachsmuth,B.D. and Cagan,R.L. (2000) The Drosophila bcl-2 family member dBorg-1 functions in the apoptotic response to UV-irradiation. Curr. Biol., 10, 547–550. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
