

Drosophila cyclin E interacts with components of the Brahma complex
- PMID: 12093739
- PMCID: PMC126084
- DOI: 10.1093/emboj/cdf334
Drosophila cyclin E interacts with components of the Brahma complex
Abstract
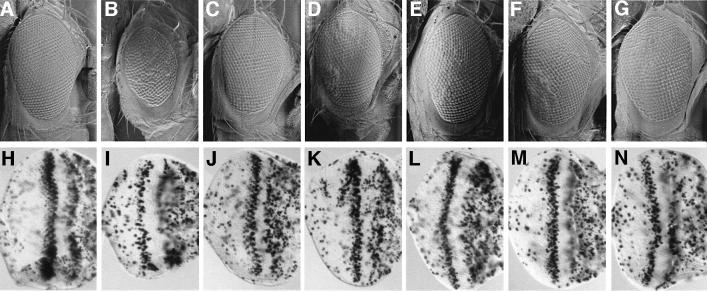
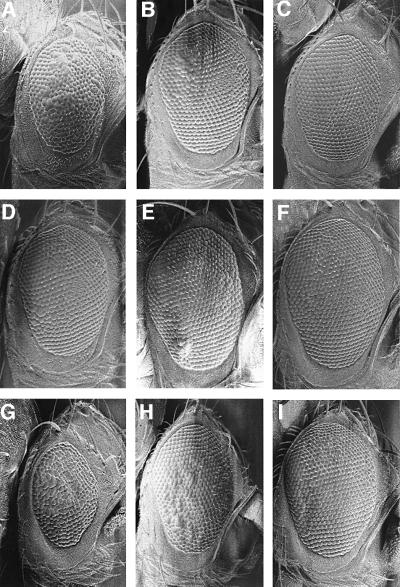
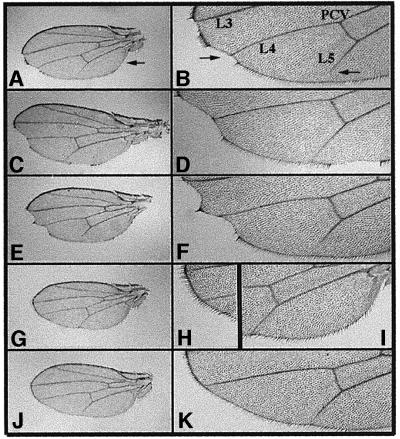
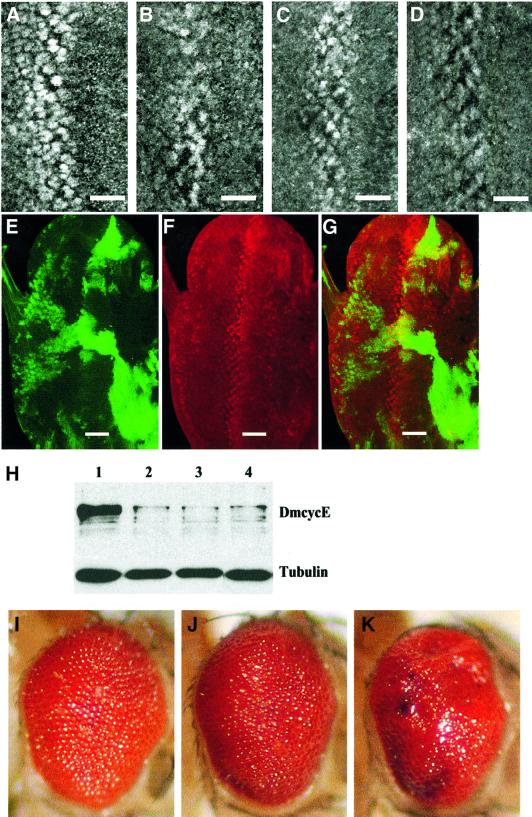
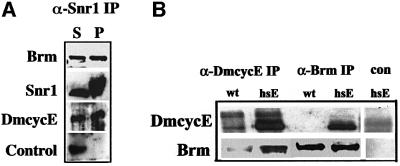
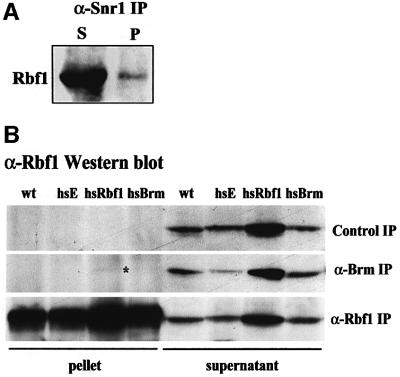
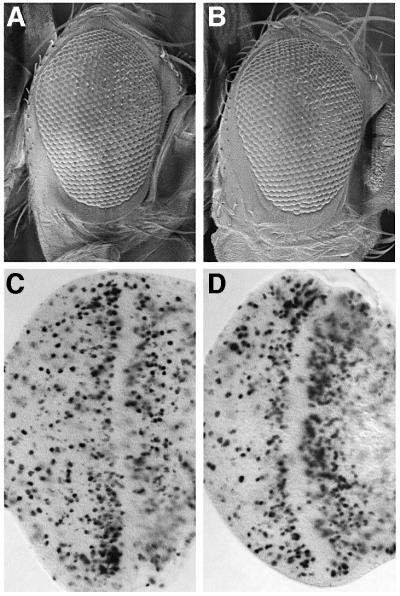
Cyclin E-Cdk2 is essential for S phase entry. To identify genes interacting with cyclin E, we carried out a genetic screen using a hypomorphic mutation of Drosophila cyclin E (DmcycE(JP)), which gives rise to adults with a rough eye phenotype. Amongst the dominant suppressors of DmcycE(JP), we identified brahma (brm) and moira (mor), which encode conserved core components of the Drosophila Brm complex that is highly related to the SWI-SNF ATP-dependent chromatin remodeling complex. Mutations in genes encoding other Brm complex components, including snr1 (BAP45), osa and deficiencies that remove BAP60 and BAP111 can also suppress the DmcycE(JP) eye phenotype. We show that Brm complex mutants suppress the DmcycE(JP) phenotype by increasing S phases without affecting DmcycE protein levels and that DmcycE physically interacts with Brm and Snr1 in vivo. These data suggest that the Brm complex inhibits S phase entry by acting downstream of DmcycE protein accumulation. The Brm complex also physically interacts weakly with Drosophila retinoblastoma (Rbf1), but no genetic interactions were detected, suggesting that the Brm complex and Rbf1 act largely independently to mediate G(1) arrest.
Figures
References
-
- Brand A.H. and Perrimon,N. (1993) Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development, 118, 401–415. - PubMed
-
- Bosco G., Du,W. and Orr-Weaver,T.L. (2001) DNA replication control through interaction of E2F-RB and the origin recognition complex. Nature Cell Biol., 3, 289–295. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
