

PA200, a nuclear proteasome activator involved in DNA repair
- PMID: 12093752
- PMCID: PMC126083
- DOI: 10.1093/emboj/cdf333
PA200, a nuclear proteasome activator involved in DNA repair
Abstract
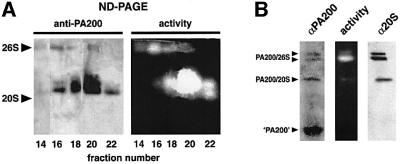
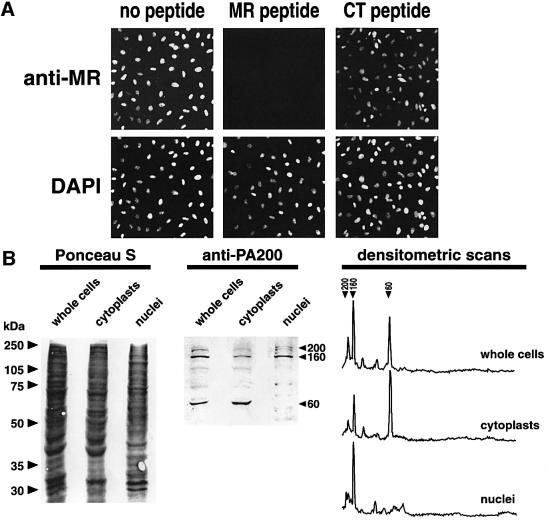
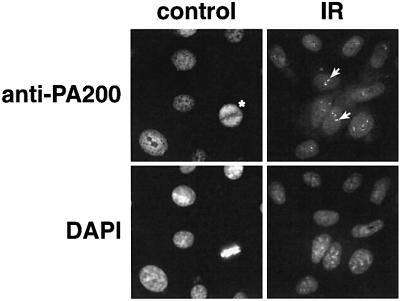
We have identified a novel 200 kDa nuclear protein that activates the proteasome. The protein, which we call PA200, has been purified to homogeneity from bovine testis and has been shown to activate proteasomal hydrolysis of peptides, but not proteins. Following gamma-irradiation of HeLa cells the uniform nuclear distribution of PA200 changes to a strikingly punctate pattern, a behavior characteristic of many DNA repair proteins. Homologs of PA200 are present in worms, plants and yeast. Others have shown that mutation of yeast PA200 results in hypersensitivity to bleomycin, and exposure of yeast to DNA damaging agents induces the PA200 message. Taken together, these findings implicate PA200 in DNA repair, possibly by recruiting proteasomes to double strand breaks.
Figures

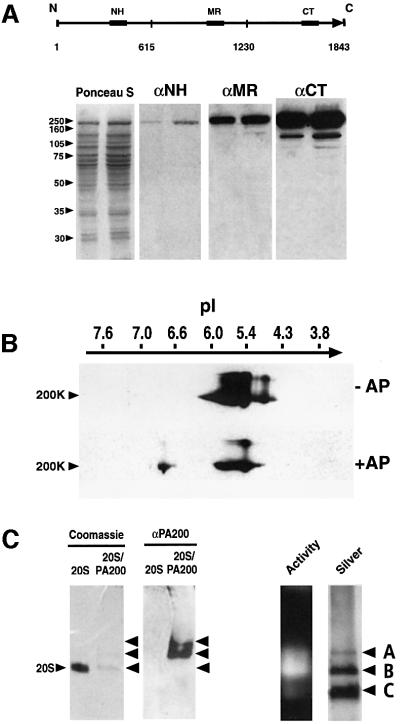
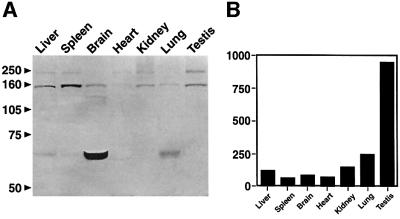
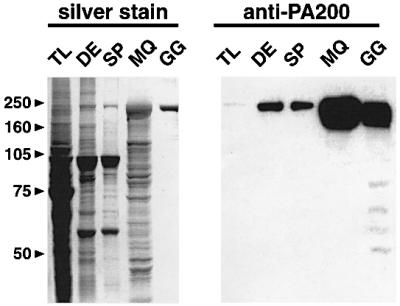
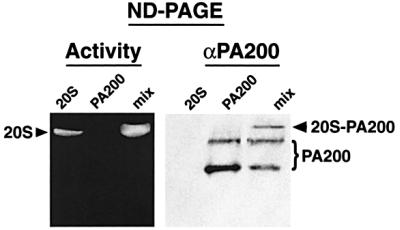
References
-
- Baumeister W., Walz,J., Zhül,F. and Seemüller,E. (1998) The proteasome: paradigm of a self-compartmentalizing protease. Cell, 92, 367–380. - PubMed
-
- Bertolaet B.L., Clarke,D.J., Wolff,M., Watson,M.H., Henze,M., Divita,G. and Reed,S.I. (2001) UBA domains mediate protein–protein interactions between two DNA damage-inducible proteins. J. Mol. Biol., 313, 955–963. - PubMed
-
- Bochtler M., Ditzel,L., Groll,M., Hartmann,C. and Huber,R. (1999) The proteasome. Annu. Rev. Biophys. Biomol. Struct., 28, 295–317. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
