

SMN, the spinal muscular atrophy protein, forms a pre-import snRNP complex with snurportin1 and importin beta
- PMID: 12095920
- PMCID: PMC1630493
- DOI: 10.1093/hmg/11.15.1785
SMN, the spinal muscular atrophy protein, forms a pre-import snRNP complex with snurportin1 and importin beta
Abstract
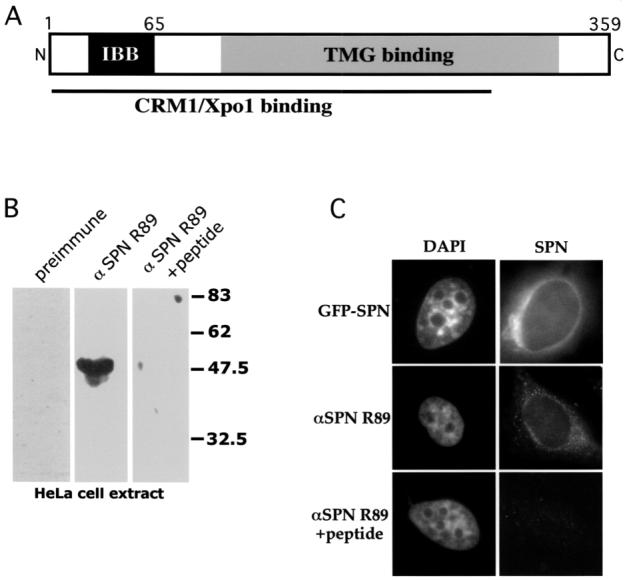
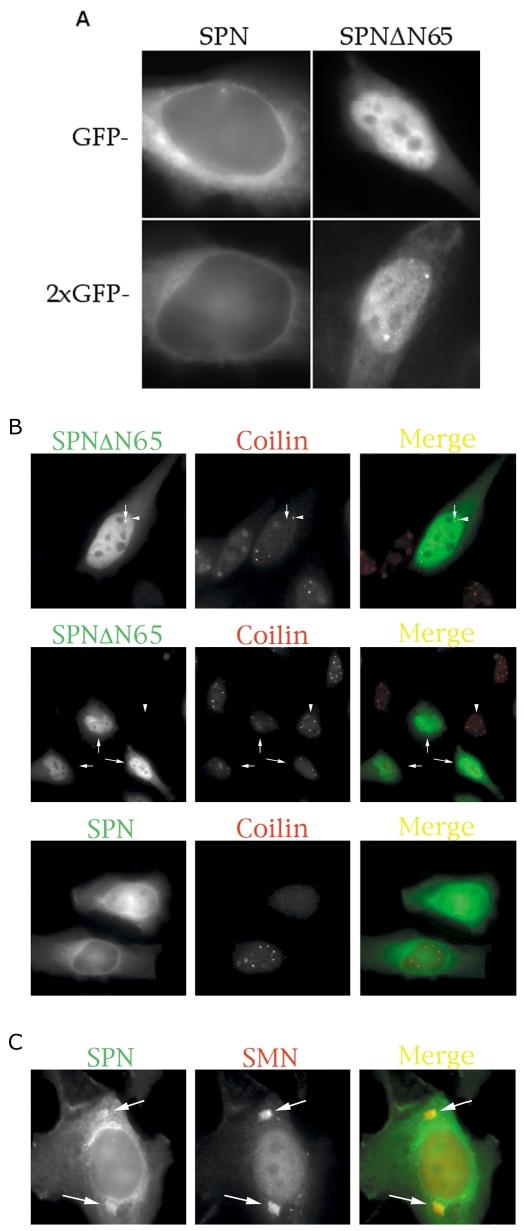
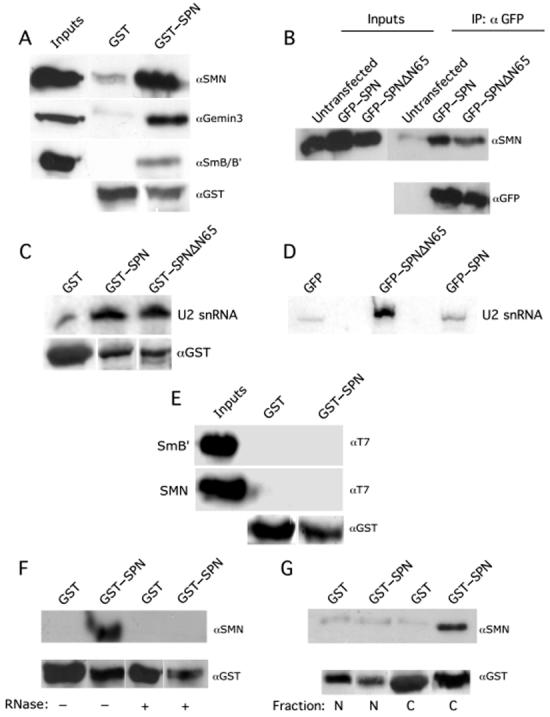
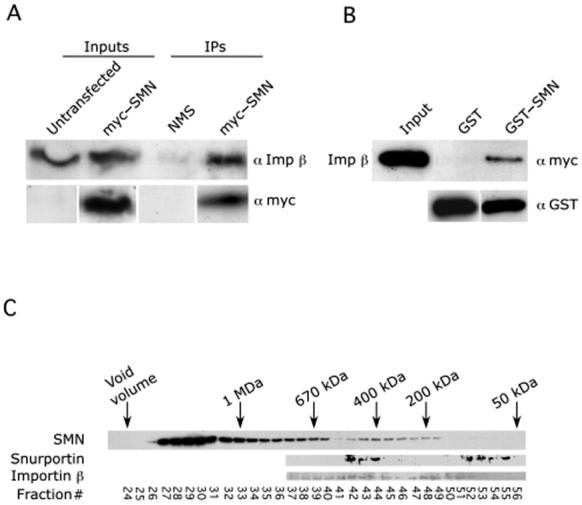
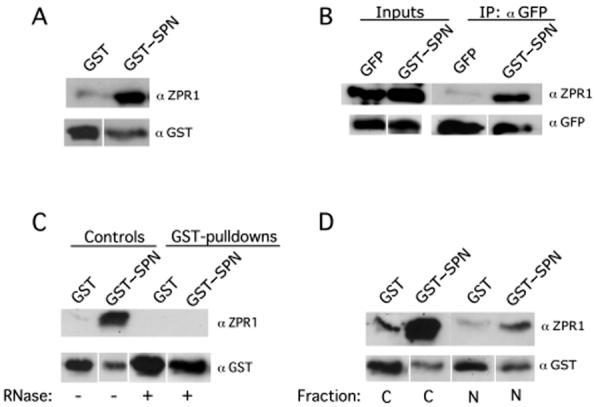
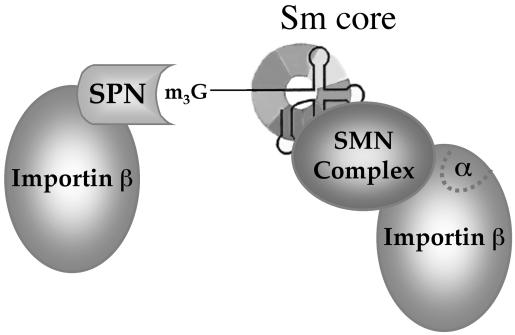
The survival of motor neuron (SMN) protein is mutated in patients with spinal muscular atrophy (SMA). SMN is part of a multiprotein complex required for biogenesis of the Sm class of small nuclear ribonucleoproteins (snRNPs). Following assembly of the Sm core domain, snRNPs are transported to the nucleus via importin beta. Sm snRNPs contain a nuclear localization signal (NLS) consisting of a 2,2,7-trimethylguanosine (TMG) cap and the Sm core. Snurportin1 (SPN) is the adaptor protein that recognizes both the TMG cap and importin beta. Here, we report that a mutant SPN construct lacking the importin beta binding domain (IBB), but containing an intact TMG cap-binding domain, localizes primarily to the nucleus, whereas full-length SPN localizes to the cytoplasm. The nuclear localization of the mutant SPN was not a result of passive diffusion through the nuclear pores. Importantly, we found that SPN interacts with SMN, Gemin3, Sm snRNPs and importin beta. In the presence of ribonucleases, the interactions with SMN and Sm proteins were abolished, indicating that snRNAs mediate this interplay. Cell fractionation studies showed that SPN binds preferentially to cytoplasmic SMN complexes. Notably, we found that SMN directly interacts with importin beta in a GST-pulldown assay, suggesting that the SMN complex might represent the Sm core NLS receptor predicted by previous studies. Therefore, we conclude that, following Sm protein assembly, the SMN complex persists until the final stages of cytoplasmic snRNP maturation and may provide somatic cell RNPs with an alternative NLS.
Figures
References
-
- Görlich D, Mattaj IW. Nucleocytoplasmic transport. Science. 1996;271:1513–1518. - PubMed
-
- Allen TD, Cronshaw JM, Bagley S, Kiseleva E, Goldberg MW. The nuclear pore complex: mediator of translocation between nucleus and cytoplasm. J. Cell Sci. 2000;113:1651–1659. - PubMed
-
- Smith AE, Slepchenko BM, Schaff JC, Loew LM, Macara IG. Systems analysis of Ran transport. Science. 2002;295:488–491. - PubMed
-
- Ryan KJ, Wente SR. The nuclear pore complex: a protein machine bridging the nucleus and cytoplasm. Curr. Opin. Cell Biol. 2000;12:361–371. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
