

KCNE4 is an inhibitory subunit to the KCNQ1 channel
- PMID: 12096056
- PMCID: PMC2290389
- DOI: 10.1113/jphysiol.2002.017301
KCNE4 is an inhibitory subunit to the KCNQ1 channel
Abstract
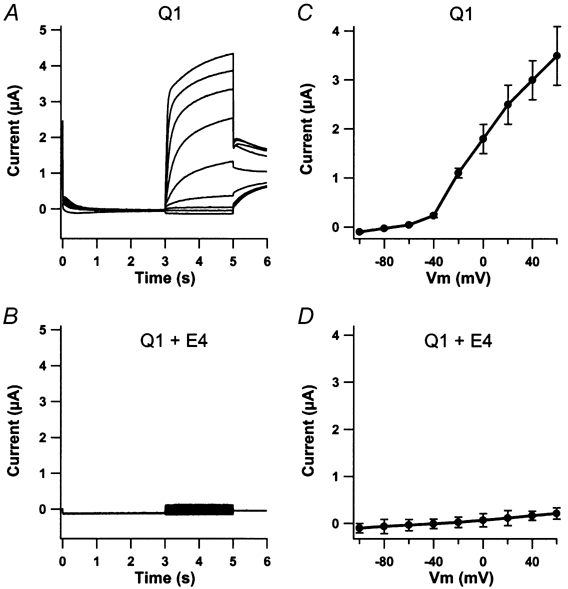
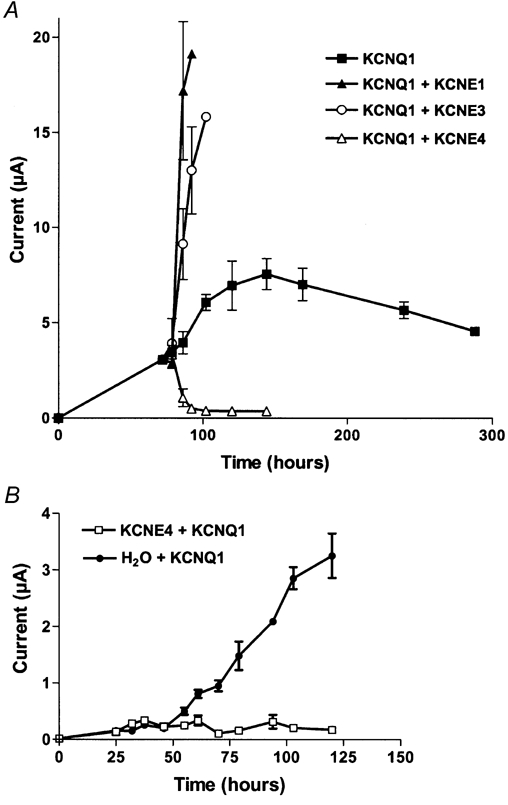
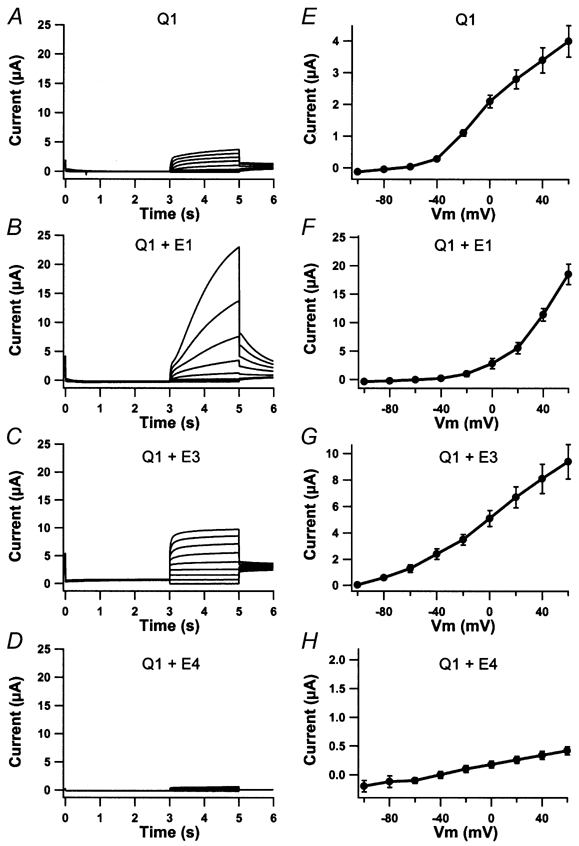
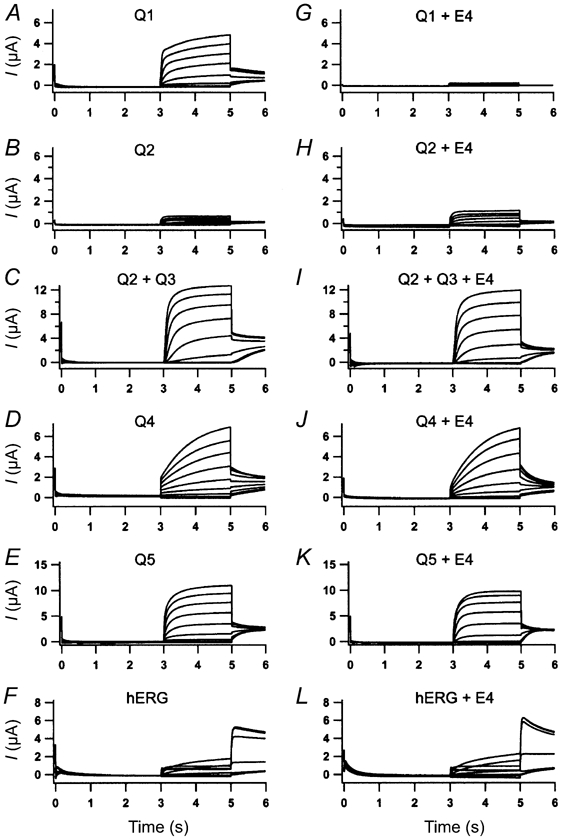
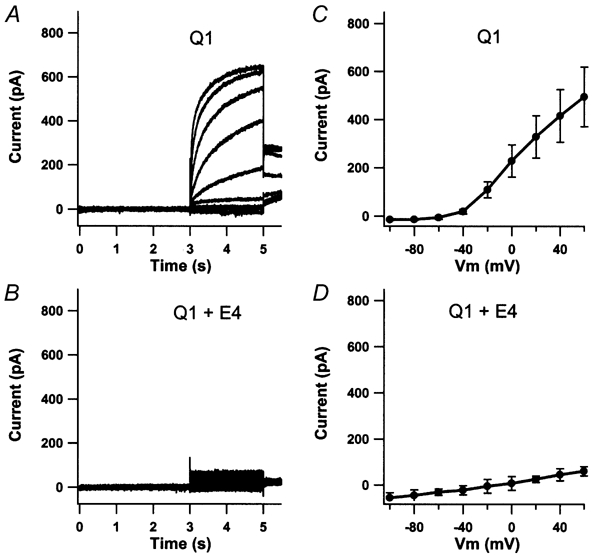
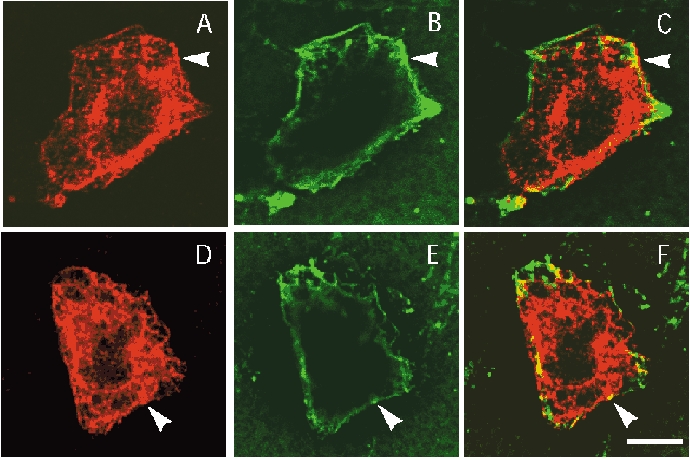
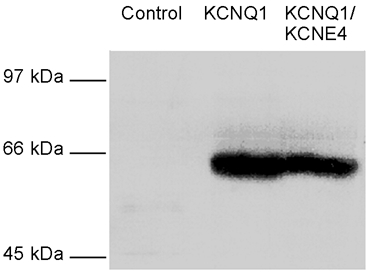
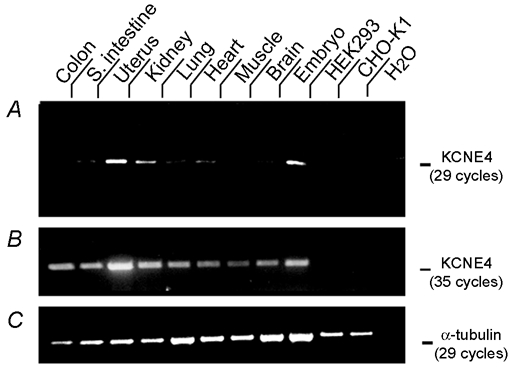
KCNE4 is a membrane protein belonging to a family of single transmembrane domain proteins known to have dramatic effect on the gating of certain potassium channels. However, no functional role of KCNE4 has been suggested so far. In the present paper we demonstrate that KCNE4 is an inhibitory subunit to KCNQ1 channels. Co-expression of KCNQ1 and KCNE4 in Xenopus oocytes completely inhibited the KCNQ1 current. This was reproduced in mammalian CHO-K1 cells. Experiments with delayed expression of mRNA coding for KCNE4 in KCNQ1-expressing oocytes suggested that KCNE4 exerts its effect on KCNQ1 channels already expressed in the plasma membrane. This notion was supported by immunocytochemical studies and Western blotting, showing no significant difference in plasma membrane expression of KCNQ1 channels in the presence or absence of KCNE4. The impact of KCNE4 on KCNQ1 was specific since no effect of KCNE4 could be detected if co-expressed with KCNQ2-5 channels or hERG1 channels. RT-PCR studies revealed high KCNE4 expression in embryos and adult uterus, where significant expression of KCNQ1 channels has also been demonstrated.
Figures
References
-
- Abbott GW, Butler MH, Bendahhou S, Dalakas MC, Ptacek LJ, Goldstein SA. MiRP2 forms potassium channels in skeletal muscle with Kv3. 4 and is associated with periodic paralysis. Cell. 2001;104:217–231. - PubMed
-
- Abbott GW, Sesti F, Splawski I, Buck ME, Lehmann MH, Timothy KW, Keating MT, Goldstein SA. MiRP1 forms IKr potassium channels with HERG and is associated with cardiac arrhythmia. Cell. 1999;97:175–187. - PubMed
-
- Barhanin J, Lesage F, Guillemare E, Fink M, Lazdunski M, Romey G. KVLQT1 and lsK (minK) proteins associate to form the I(Ks) cardiac potassium current. Nature. 1996;384:78–80. - PubMed
-
- Boucherot A, Schreiber R, Kunzelmann K. Regulation and properties of KCNQ1 (KVLQT1) and impact of the cystic fibrosis transmembrane conductance regulator. Journal of Membrane Biology. 2001;182:39–47. - PubMed
-
- Demolombe S, Franco D, de Boer P, Kuperschmidt S, Roden D, Pereon Y, Jarry A, Moorman AF, Escande D. Differential expression of KvLQT1 and its regulator IsK in mouse epithelia. American Journal of Physiology – Cell Physiology. 2001;280:C359–372. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases