

Control of feedforward dendritic inhibition by NMDA receptor-dependent spike timing in hippocampal interneurons
- PMID: 12097498
- PMCID: PMC6758215
- DOI: 10.1523/JNEUROSCI.22-13-05462.2002
Control of feedforward dendritic inhibition by NMDA receptor-dependent spike timing in hippocampal interneurons
Abstract
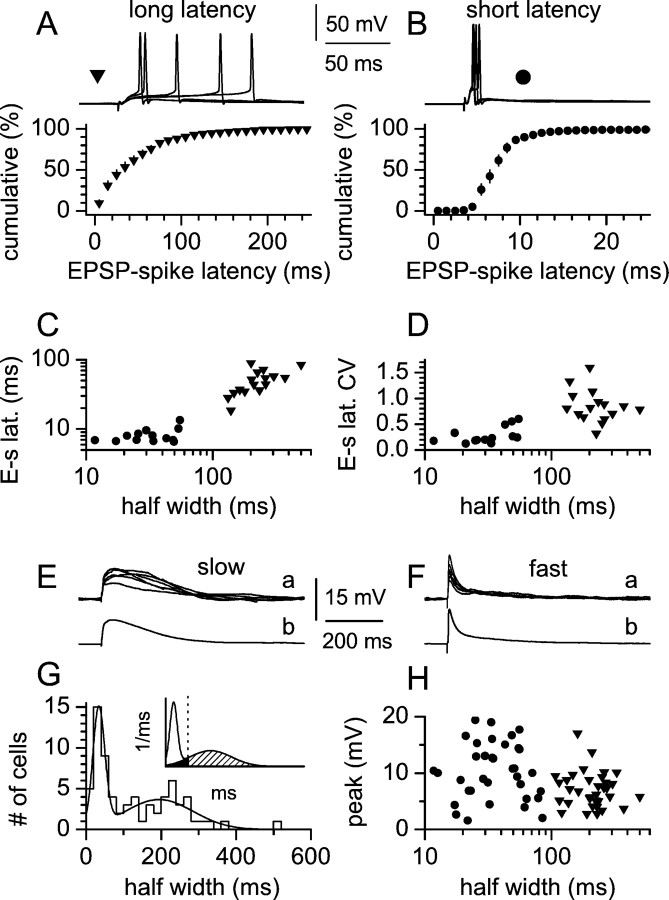
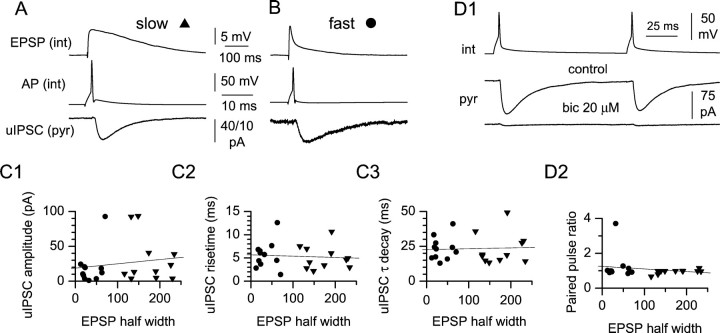
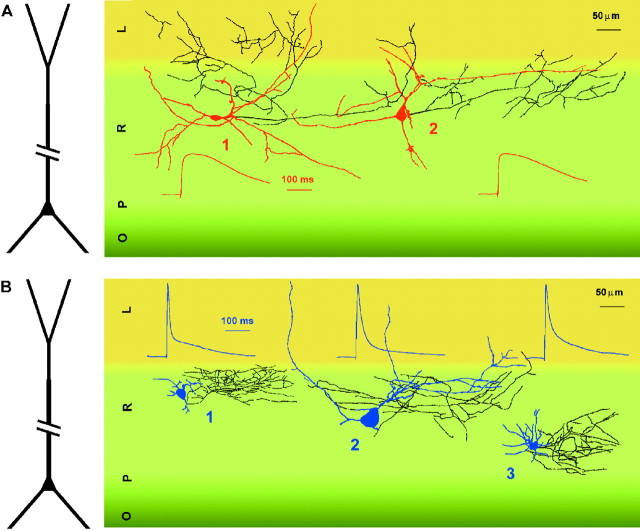
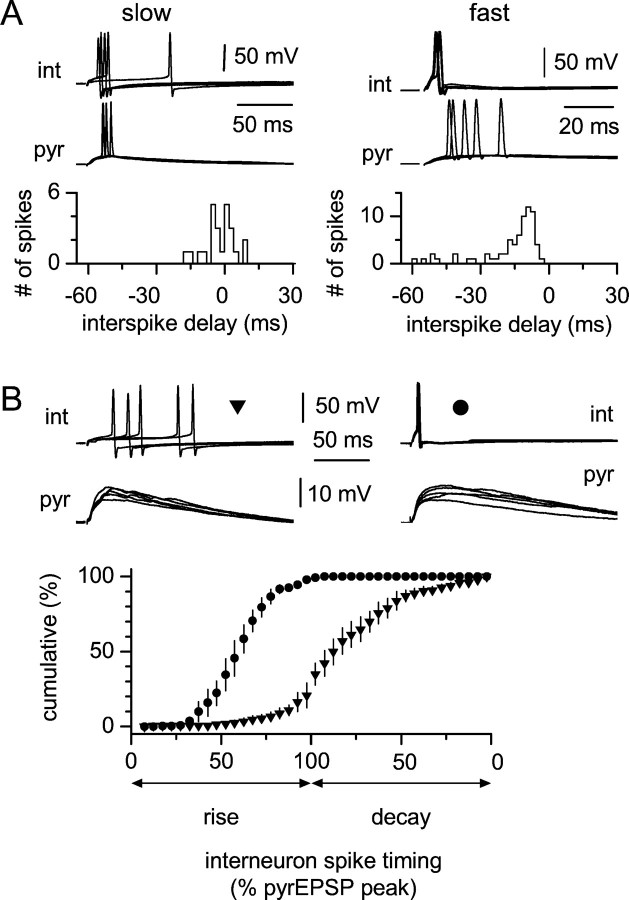
Two putative functional populations of feedforward interneurons with distinct spike-timing properties were identified in stratum radiatum of the CA1 rat hippocampus. Interneurons with fast (half width, <100 msec) EPSPs fired after short EPSP-spike latencies and with a high degree of temporal precision compared with cells with slow (half width, >100 msec) EPSPs. Spike timing in fast and slow interneurons occurred at different phases of the EPSPs of simultaneously activated pyramidal cells. In addition, firing of fast interneurons preceded action potentials in principal neurons, whereas action potentials in slow interneurons could either precede or follow firing in pyramidal cells. Temporal integration of separate inputs leading to synaptically evoked firing was more prominent in slow than fast interneurons. Functional diversity between the two putative populations was abolished by the NMDA receptor (NMDAR) antagonist d-(-)-2-amino-5-phosphonopentanoic acid (d-AP-5). The axon of both cell types was primarily restricted to striatum radiatum or to striatum lacunosum-moleculare in the case of slow cells, suggesting targeting of principal cell dendrites for the majority of the cells of this study. Indeed, firing of slow and fast interneurons generated similar unitary IPSCs (uIPSCs) in pyramidal neurons. uIPSCs were mediated by GABA(A) receptors and had in most cases small amplitudes and slow kinetics. Our results suggest that functionally heterogeneous interneurons encode the temporal properties of dendritic feedforward inhibition, and that NMDARs play an essential role in shaping the integrative properties of interneurons and in setting the timing of GABA release.
Figures



Similar articles
-
Membrane properties and synaptic responses of interneurons located near the stratum lacunosum-moleculare/radiatum border of area CA1 in whole-cell recordings from rat hippocampal slices.J Neurophysiol. 1994 Jun;71(6):2217-35. doi: 10.1152/jn.1994.71.6.2217. J Neurophysiol. 1994. PMID: 7931512
-
Membrane properties and synaptic currents evoked in CA1 interneuron subtypes in rat hippocampal slices.J Neurophysiol. 1996 Jul;76(1):1-16. doi: 10.1152/jn.1996.76.1.1. J Neurophysiol. 1996. PMID: 8836204
-
Factors determining the efficacy of distal excitatory synapses in rat hippocampal CA1 pyramidal neurones.J Physiol. 1998 Mar 1;507 ( Pt 2)(Pt 2):441-62. doi: 10.1111/j.1469-7793.1998.441bt.x. J Physiol. 1998. PMID: 9518704 Free PMC article.
-
GABAergic interneurons targeting dendrites of pyramidal cells in the CA1 area of the hippocampus.Eur J Neurosci. 2009 Sep;30(6):947-57. doi: 10.1111/j.1460-9568.2009.06913.x. Epub 2009 Sep 4. Eur J Neurosci. 2009. PMID: 19735288 Review.
-
Neurogliaform cells and other interneurons of stratum lacunosum-moleculare gate entorhinal-hippocampal dialogue.J Physiol. 2011 Apr 15;589(Pt 8):1875-83. doi: 10.1113/jphysiol.2010.201004. Epub 2010 Dec 6. J Physiol. 2011. PMID: 21135049 Free PMC article. Review.
Cited by
-
Astrocyte glutamate transporters regulate metabotropic glutamate receptor-mediated excitation of hippocampal interneurons.J Neurosci. 2004 May 12;24(19):4551-9. doi: 10.1523/JNEUROSCI.5217-03.2004. J Neurosci. 2004. PMID: 15140926 Free PMC article.
-
Electrical coupling between interneurons with different excitable properties in the stratum lacunosum-moleculare of the juvenile CA1 rat hippocampus.J Neurosci. 2005 Sep 21;25(38):8686-95. doi: 10.1523/JNEUROSCI.2810-05.2005. J Neurosci. 2005. PMID: 16177037 Free PMC article.
-
New approaches to the management of schizophrenia: focus on aberrant hippocampal drive of dopamine pathways.Drug Des Devel Ther. 2014 Jul 2;8:887-96. doi: 10.2147/DDDT.S42708. eCollection 2014. Drug Des Devel Ther. 2014. PMID: 25061280 Free PMC article. Review.
-
Membrane Potential Dynamics of CA1 Pyramidal Neurons during Hippocampal Ripples in Awake Mice.Neuron. 2016 Feb 17;89(4):800-13. doi: 10.1016/j.neuron.2016.01.014. Neuron. 2016. PMID: 26889811 Free PMC article.
-
The role of glutamatergic inputs onto parvalbumin-positive interneurons: relevance for schizophrenia.Rev Neurosci. 2012 Jan 9;23(1):97-109. doi: 10.1515/revneuro-2011-0059. Rev Neurosci. 2012. PMID: 22718616 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous