

Thermotaxis in Caenorhabditis elegans analyzed by measuring responses to defined Thermal stimuli
- PMID: 12097525
- PMCID: PMC6758190
- DOI: 10.1523/JNEUROSCI.22-13-05727.2002
Thermotaxis in Caenorhabditis elegans analyzed by measuring responses to defined Thermal stimuli
Abstract
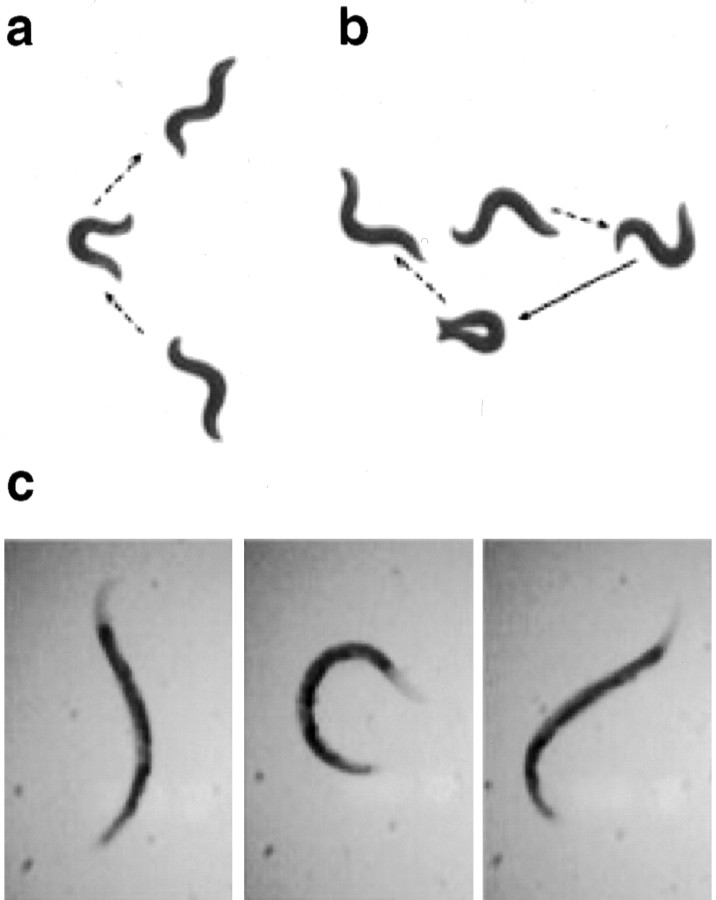
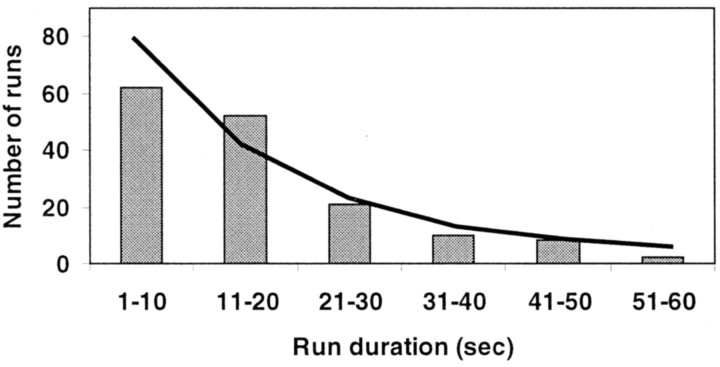
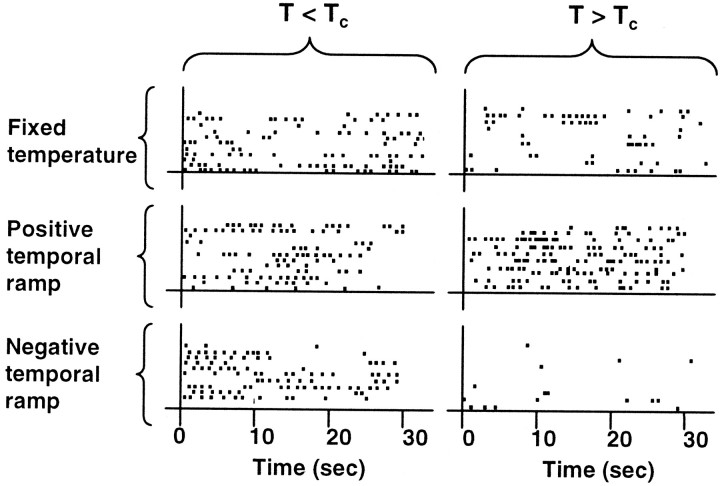
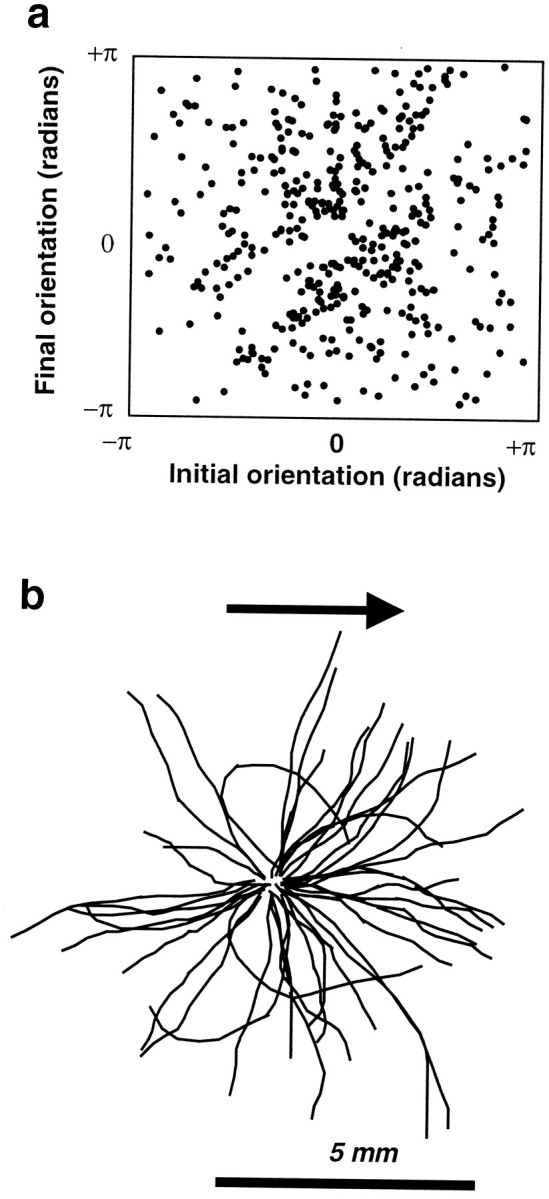
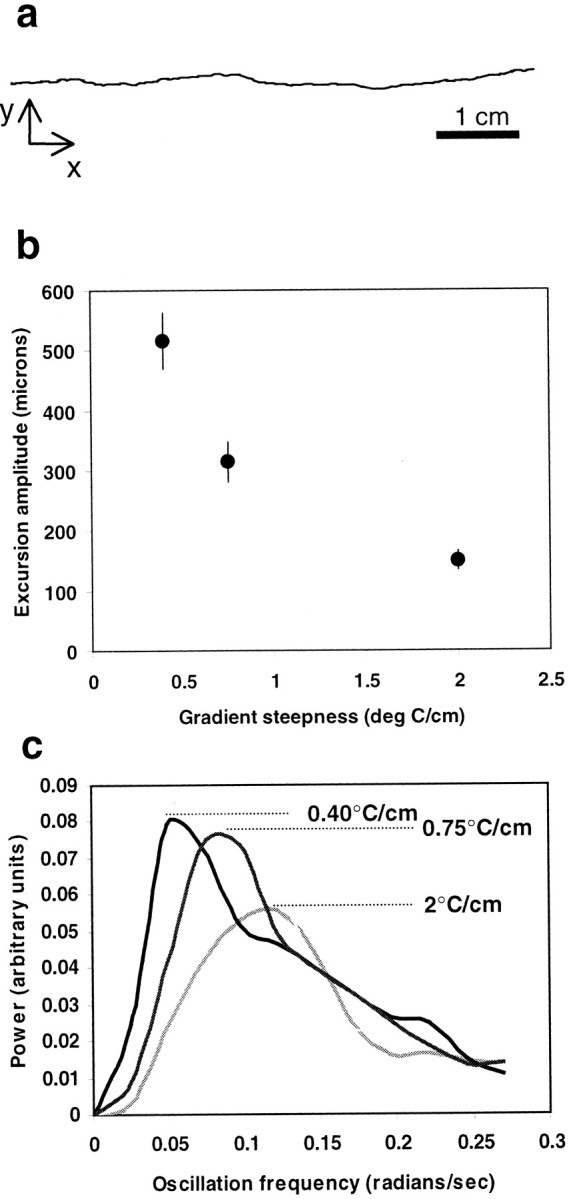
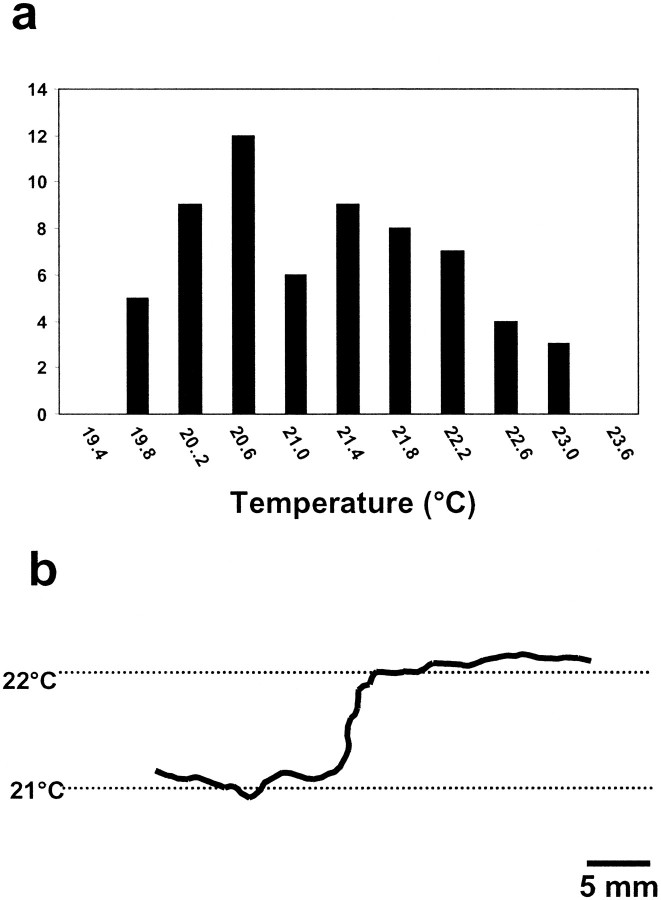
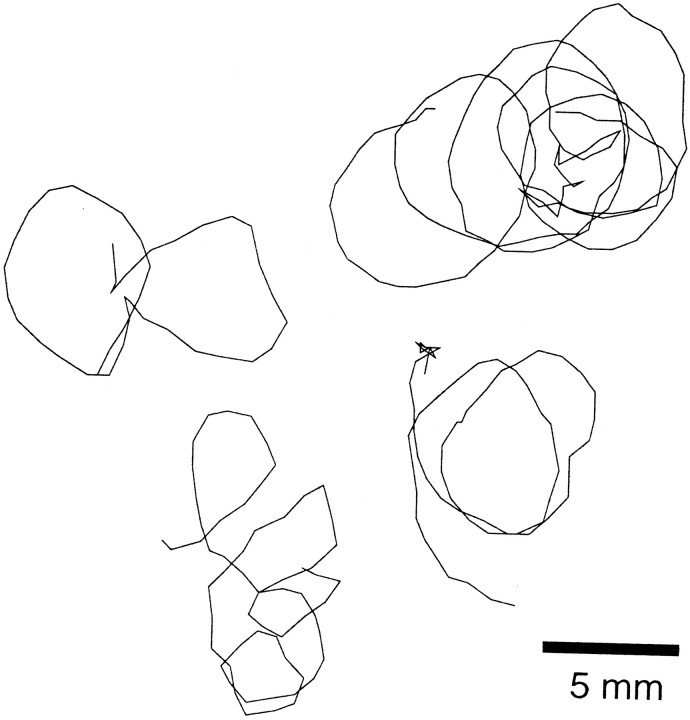
In a spatial thermal gradient, Caenorhabditis elegans migrates toward and then isothermally tracks near its cultivation temperature. A current model for thermotactic behavior involves a thermophilic drive (involving the neurons AFD and AIY) and cryophilic drive (involving the neuron AIZ) that balance at the cultivation temperature. Here, we analyze the movements of individual worms responding to defined thermal gradients. We found evidence for a mechanism for migration down thermal gradients that is active at temperatures above the cultivation temperature, and a mechanism for isothermal tracking that is active near the cultivation temperature. However, we found no evidence for a mechanism for migration up thermal gradients at temperatures below the cultivation temperature that might have supported the model of opposing drives. The mechanisms for migration down gradients and isothermal tracking control the worm's movements in different manners. Migration down gradients works by shortening (lengthening) the duration of forward movement in response to positive (negative) temperature changes. Isothermal tracking works by orienting persistent forward movement to offset temperature changes. We believe preference for the cultivation temperature is not at the balance between two drives. Instead, the worm activates the mechanism for isothermal tracking near the cultivation temperature and inactivates the mechanism for migration down gradients near or below the cultivation temperature. Inactivation of the mechanism for migration down gradients near or below the cultivation temperature requires the neurons AFD and AIY.
Figures
References
-
- Croll NA. Components and patterns in the behavior of the nematode Caenorhabditis elegans. J Zool Lond. 1975;16:159–176.
-
- Gomez M, DeCastro E, Guarin E, Sasakura H, Kujara A, Mori I, Bartfai T, Bargmann CI, Nef P. Ca2+ signaling via the neuronal calcium sensor ncs-1 regulates associative learning and memory in C. elegans. Neuron. 2001;30:241–248. - PubMed
-
- Hobert O, Mori I, Yamashita Y, Honda H, Ohshima Y, Liu Y, Ruvkun G. Regulation of interneuron function in the C. elegans thermoregulatory pathway by the ttx-3 LIM homeobox gene. Neuron. 1997;19:345–357. - PubMed
-
- Mori I. Genetics of chemotaxis and thermotaxis in the nematode Caenorhabditis elegans. Annu Rev Genet. 1999;33:399–422. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources