

RNA editing in hepatitis delta virus genotype III requires a branched double-hairpin RNA structure
- PMID: 12097551
- PMCID: PMC136351
- DOI: 10.1128/jvi.76.15.7385-7397.2002
RNA editing in hepatitis delta virus genotype III requires a branched double-hairpin RNA structure
Abstract
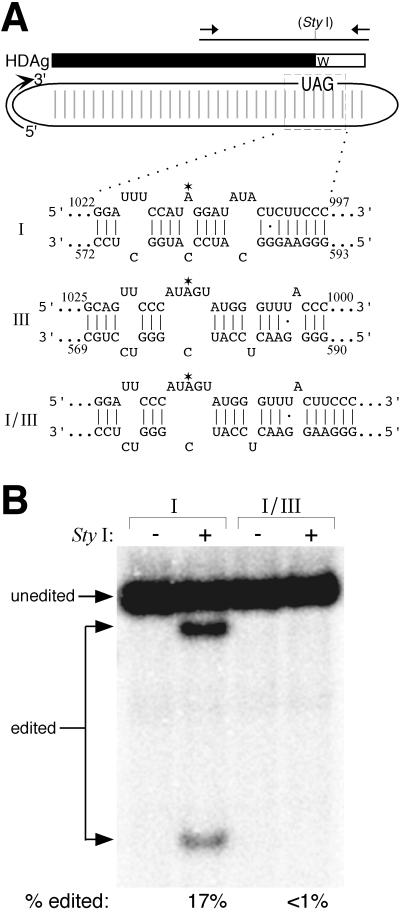
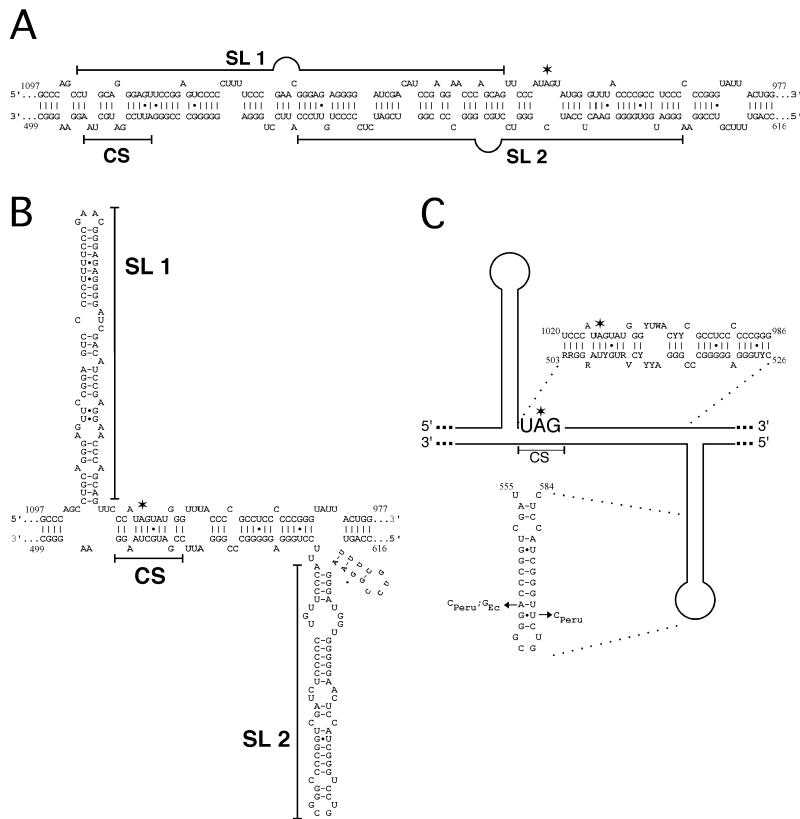
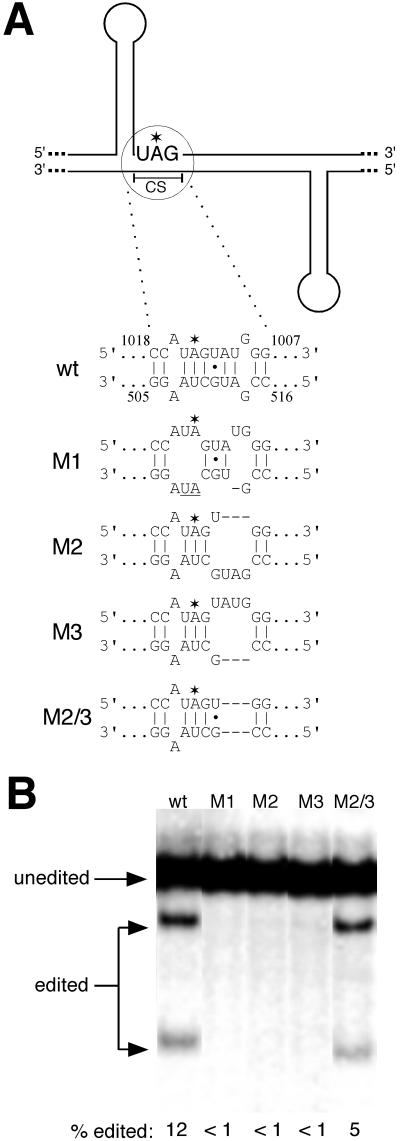
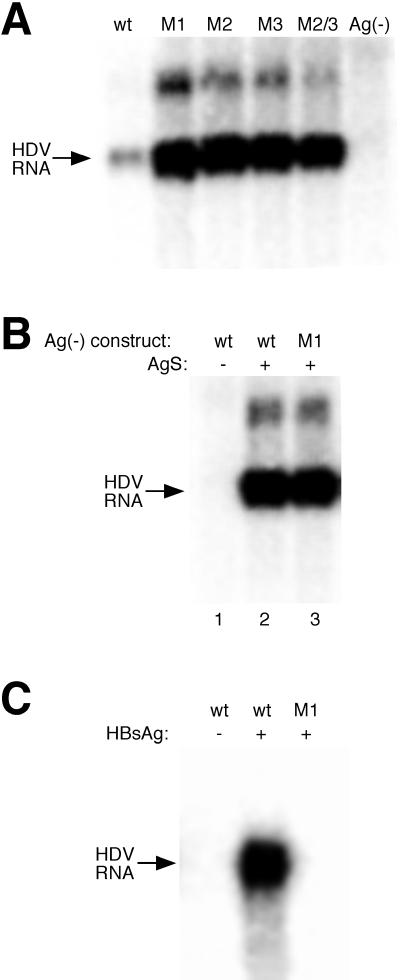
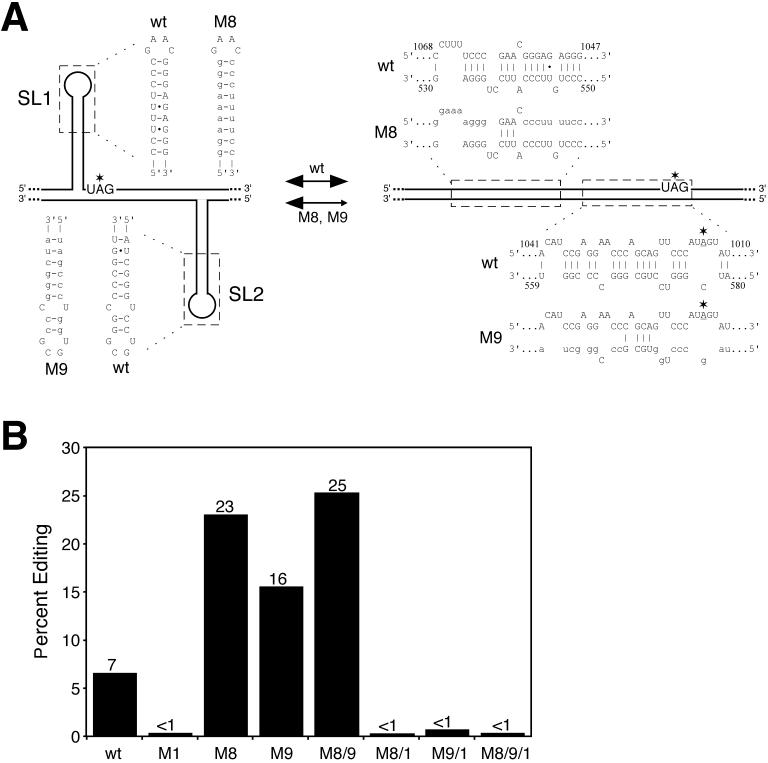
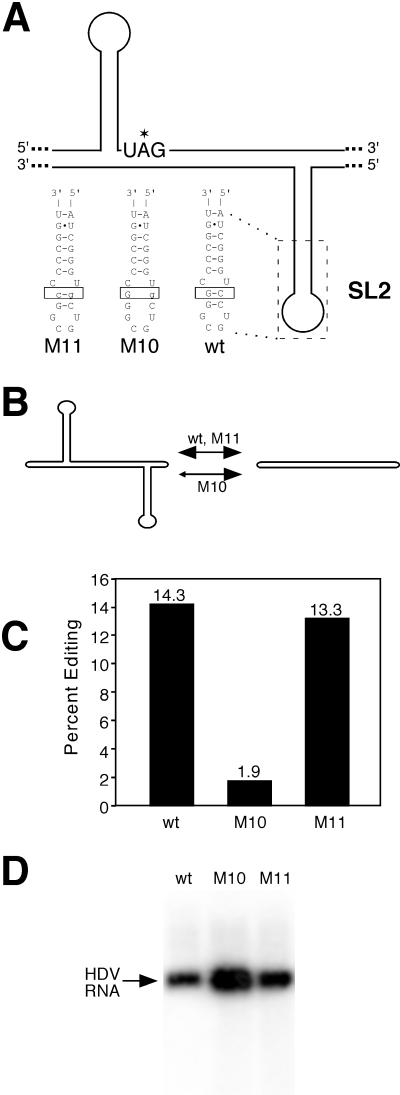
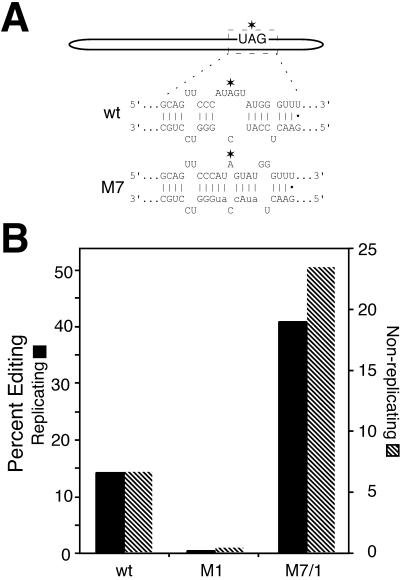
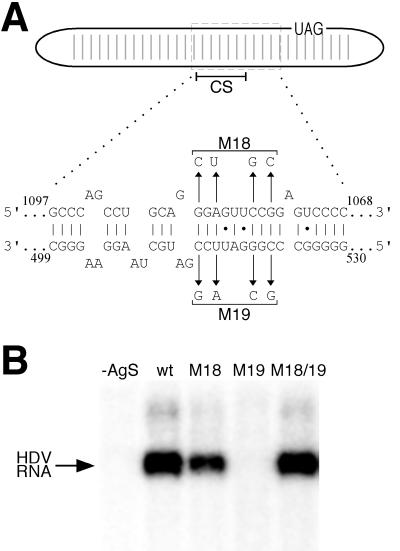
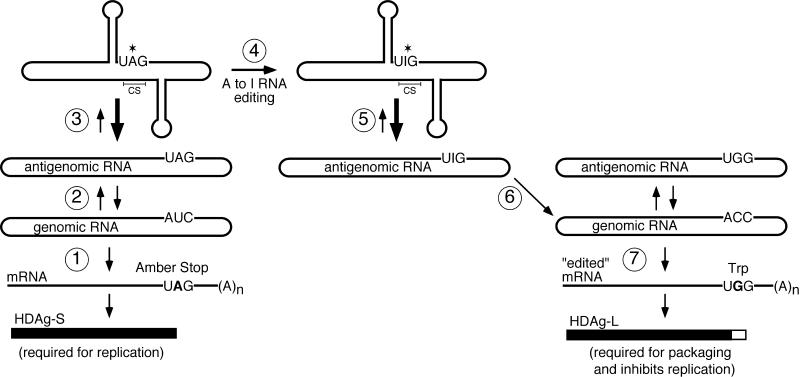
RNA editing at the amber/W site plays a central role in the replication scheme of hepatitis delta virus (HDV), allowing the virus to produce two functionally distinct forms of the sole viral protein, hepatitis delta antigen (HDAg), from the same open reading frame. Editing is carried out by a cellular activity known as ADAR (adenosine deaminase), which acts on RNA substrates that are at least partially double stranded. In HDV genotype I, editing requires a highly conserved base-paired structure that occurs within the context of the unbranched rod structure characteristic of HDV RNA. This base-paired structure is disrupted in the unbranched rod of HDV genotype III, which is the most distantly related of the three known HDV genotypes and is associated with the most severe disease. Here I show that RNA editing in HDV genotype III requires a branched double-hairpin structure that deviates substantially from the unbranched rod structure, involving the rearrangement of nearly 80 bp. The structure includes a UNCG RNA tetraloop, a highly stable structural motif frequently involved in the folding of large RNAs such as rRNA. The double-hairpin structure is required for editing, and hence for virion formation, but not for HDV RNA replication, which requires the unbranched rod structure. HDV genotype III thus relies on a dynamic conformational switch between the two different RNA structures: the unbranched rod characteristic of HDV RNA and a branched double-hairpin structure that is required for RNA editing. The different mechanisms of editing in genotypes I and III underscore their functional differences and may be related to pathogenic differences as well.
Figures
References
-
- Abdelkafi, M., N. Leulliot, V. Baumruk, L. Bednarova, P. Y. Turpin, A. Namane, C. Gouyette, T. Huynh-Dinh, and M. Ghomi. 1998. Structural features of the UCCG and UGCG tetraloops in very short hairpins as evidenced by optical spectroscopy. Biochemistry 37:7878-7884. - PubMed
-
- Been, M. D., and G. S. Wickham. 1997. Self-cleaving ribozymes of hepatitis delta virus RNA. Eur. J. Biochem. 247:741-753. - PubMed
-
- Bergmann, K. F., and J. L. Gerin. 1986. Antigens of hepatitis delta virus in the liver and serum of humans and animals. J. Infect. Dis. 154:702-706. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
