

doi: 10.1073/pnas.152002699.
Epub 2002 Jul 3.
Bradykinin-12-lipoxygenase-VR1 signaling pathway for inflammatory hyperalgesia
Affiliations
- PMID: 12097645
- PMCID: PMC126639
- DOI: 10.1073/pnas.152002699
Item in Clipboard
Bradykinin-12-lipoxygenase-VR1 signaling pathway for inflammatory hyperalgesia
Proc Natl Acad Sci U S A.
.
Abstract
The capsaicin-sensitive vanilloid receptor (VR1) was recently shown to play an important role in inflammatory pain (hyperalgesia), but the underlying mechanism is unknown. We hypothesized that pain-producing inflammatory mediators activate capsaicin receptors by inducing the production of fatty acid agonists of VR1. This study demonstrates that bradykinin, acting at B2 bradykinin receptors, excites sensory nerve endings by activating capsaicin receptors via production of 12-lipoxygenase metabolites of arachidonic acid. This finding identifies a mechanism that might be targeted in the development of new therapeutic strategies for the treatment of inflammatory pain.
Figures
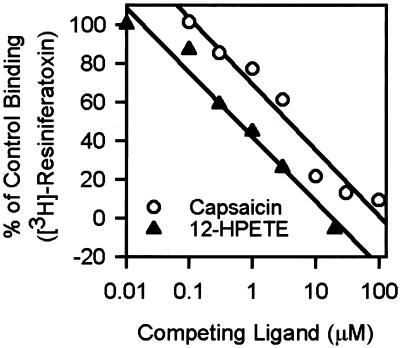
12-HPETE inhibits the specific binding of [3H]-RTX to VR1-HEK cells. Cells were incubated with various concentrations of 12-HPETE and 100 pM [3H]-RTX. Capsaicin was used as a control. Apparent Ki values for 12-HPETE and capsaicin were 0.35 and 2.5 μM, respectively (quadruplicate preparations).
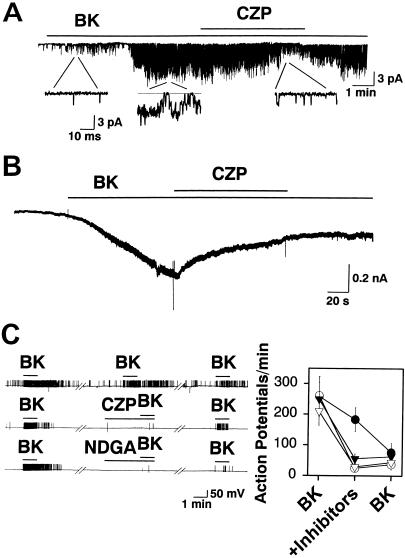
BK excites cultured sensory neurons via the PLA2/lipoxygenase/VR1 pathway. (A) Bath application of 10 μM CZP antagonizes single-channel currents activated by bath application of 1 μM BK in a cell-attached patch. (B) CZP (10 μM) antagonizes whole-cell currents induced by 1 μM BK. (C) Action potential firing induced by 0.1 μM BK is antagonized by 5 μM CZP (open circles, n = 9), a lipoxygenase inhibitor, 5 μM NDGA (open triangles, n = 9), and a PLA2 inhibitor, 10 μM QN (filled triangles, n = 6). For control experiments (filled circles, n = 10), application of BK was repeated three times. Gaps represent 5-min intervals.
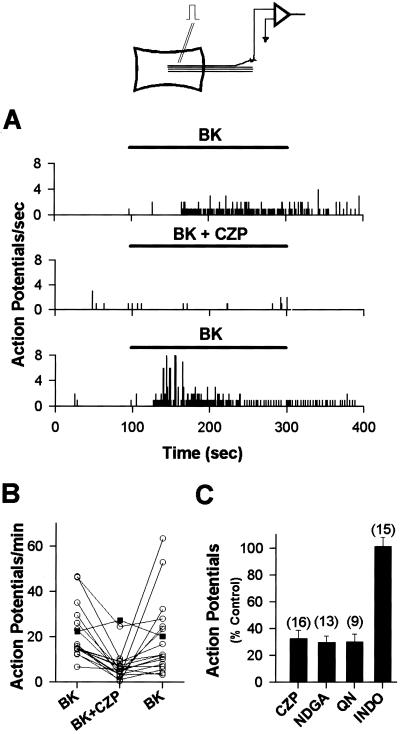
BK (1 μM)-induced excitation of cutaneous nerve fibers in the in vitro skin-nerve preparation is antagonized by 10 μM CZP, 20 μM NDGA, and 10 μM QN, but not by a cyclooxygenase inhibitor, 2 μM INDO. (A) Peristimulus-time histograms of BK-induced activity, which is reversibly inhibited by 10 μM CZP. Bin width = 100 ms. (B) CZP (10 μM) reversibly antagonizes nerve activity induced by 1 μM BK in 16 fibers (circles). Vehicle (filled squares) does not antagonize the activity (n = 7). (C) Antagonists of the PLA2/lipoxygenase/VR1 cascade significantly (P < 0.01) reduce the number of action potentials induced by 1 μM BK compared with the control response to BK before application of antagonists. INDO has no significant effect on the response to BK. Numbers in parentheses represent the number of experiments.
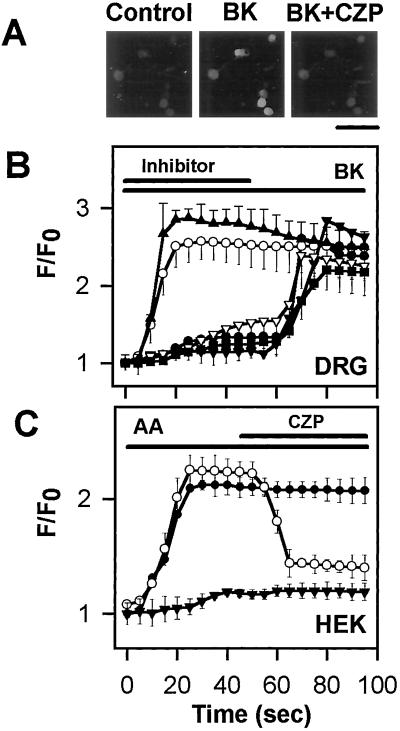
BK-evoked calcium influx is mediated by the PLA2/12–lipoxygenase/VR1 pathway. (A) Fluorescence photomicrographs show that 1 μM BK-induced [Ca2+]i increase in cultured sensory neurons is blocked by 5 μM CZP. Control represents a fluorescence photomicrograph of cultured sensory neurons preincubated with Fluo-3/AM before BK and BK and CZP (BK+CZP) application. (Bar = 100 μm.) (B) Increase in [Ca2+]i induced by 1 μM BK in sensory neurons (filled triangles, n = 15) and inhibition by an inhibitor of PLA2 (QN, open triangles), inhibitors of 12-lipoxygenase (NDGA, filled inverted triangles and baicalein, filled circles), and CZP (filled squares). A cyclooxygenase inhibitor, INDO (open circles) fails to inhibit the BK-induced increase in [Ca2+]i. (C) The 12-lipoxygenase/VR1 cascade can be reconstituted in HEK cells. AA (10 μM) evokes a large [Ca2+]i increase in HEK cells transfected with both 12-lipoxygenase and VR1 (filled circles, n = 72), but not in cells transfected with cyclooxygenase-2 and VR1 (filled triangles, n = 54). The AA-induced [Ca2+]i increase in HEK cells transfected with 12-lipoxygenase and VR1 (open circles, n = 67) is blocked by 10 μM CZP.
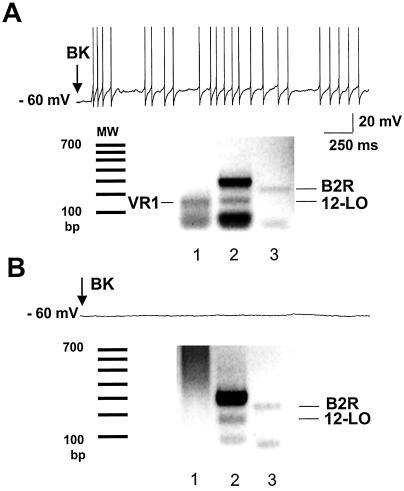
B2 BK receptor, VR1, and 12-lipoxygenase are coexpressed in most BK-responsive sensory neurons. (A) Single-cell RT-PCR products of VR1 (lane 1), 12-lipoxygenase (12-LO, lane 2), and B2 receptor (B2R, lane 3) are coexpressed in a single sensory neuron that fires action potentials (upper trace) in response to BK. MW denotes the molecular weight standards. (B) In a cell that does not respond to BK (upper trace), RT-PCR products of 12–lipoxygenase and B2R are detected, but VR1 is undetectable (lower trace).
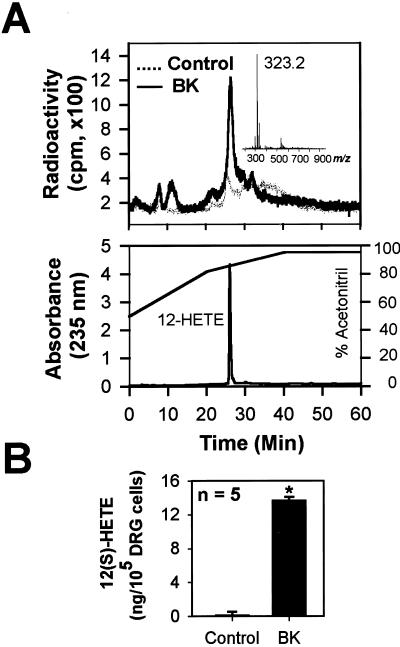
BK causes the production of 12-lipoxygenase metabolites of AA in sensory neurons. (A) Application of 1 μM BK to cultured sensory neurons preincubated with 14C-AA evokes a radio-labeled peak that is identical to 12-HETE in its retention time when detected with a high-performance liquid chromatography at 235-nm wavelength (Lower). A mass spectral analysis of the radio-labeled peak (Inset) further confirms the molecular ion peak (m/z 323.5 [M+H]+) of 14C-12-HETE (molecular weight = 322.5). Concentration of acetonitrile in the mobile solution is indicated as a solid line. (B) 12(S)-HETE specific enzyme immunoassay also detects the increase in 12(S)-HETE after BK application in sensory neurons (*, P < 0.0001).
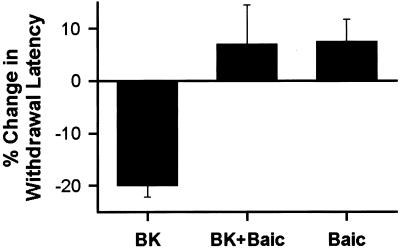
BK-induced thermal hyperalgesia requires 12-lipoxygenase activity. Injection of 1 μg of BK in the hind paw reduces the latency of the heat-evoked withdrawal reflex (n = 8). Coadministration of 1 μg of baicalein (BK + Baic) prevented the effect of BK (n = 8). Baicalein (Baic) alone had no significant effect on the paw withdrawal latency (n = 4).
References
-
- Caterina M J, Schumacher M A, Tominaga M, Rosen T A, Levine J D, Julius D. Nature (London) 1997;389:816–824. - PubMed
-
- Tominaga M, Caterina M J, Malmberg A B, Rosen T A, Gilbert H, Skinner K, Raumann B E, Basbaum A I, Julius D. Neuron. 1998;21:531–543. - PubMed
-
- Caterina M J, Leffler A, Malmberg A B, Martin W J, Trafton J, Petersen-Zeitz K R, Koltzenburg M, Basbaum A I, Julius D. Science. 2000;288:306–313. - PubMed
-
- Davis J B, Gray J, Gunthorpe M J, Hatcher J P, Davey P T, Overend P, Harries M H, Latcham J, Clapham C, Atkinson K, et al. Nature (London) 2000;405:183–187. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
