

Herpes simplex virus 1 infected neuronal and skin cells differ in their susceptibility to complement attack
- PMID: 12100729
- PMCID: PMC1782739
- DOI: 10.1046/j.1365-2567.2002.01421.x
Herpes simplex virus 1 infected neuronal and skin cells differ in their susceptibility to complement attack
Abstract
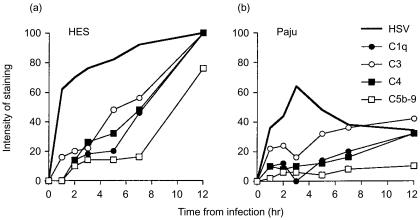
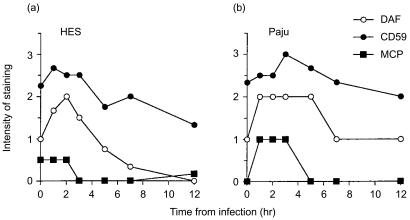
Herpes simplex virus type 1 (HSV-1) infection in neurons is lifelong and generally asymptomatic. Reactivation of this latent infection results in skin blistering whereas the respective peripheral neurons are rarely affected. Why the neuronal cells are spared while the skin cells are sacrificed is not well understood. In the present study our aim was to study whether neuronal and skin cells differ in their ability to control complement attack during HSV-1 infection. Human embryonal skin (HES) cells and neuronal Paju cells were infected by HSV-1 in vitro. Both types of infected cells activated complement but were initially resistant to membrane attack complex (MAC) deposition. During the first hours of infection the expression of the endogenous complement regulators decay accelerating factor (DAF) and CD59 increased on both HES and Paju cells. By 12 hr the infected HES cells had lost their ability to control complement attack. The expression of DAF and CD59 decreased and the cells became targets for MAC attack. In contrast, complement regulator expression on the Paju cells did not decrease below the initial level and complement C5b-9 deposition was found only on 10% of the Paju cells at 12 hr. The results suggest that HSV-infected neuronal cells are better than skin cells in protecting themselves against complement attack. This may contribute to the persistence of a latent HSV-1 infection in neuronal cells for prolonged periods.
Figures
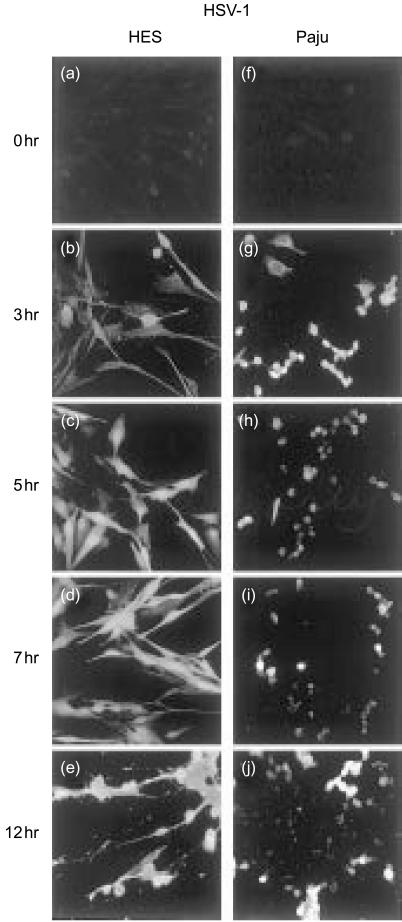
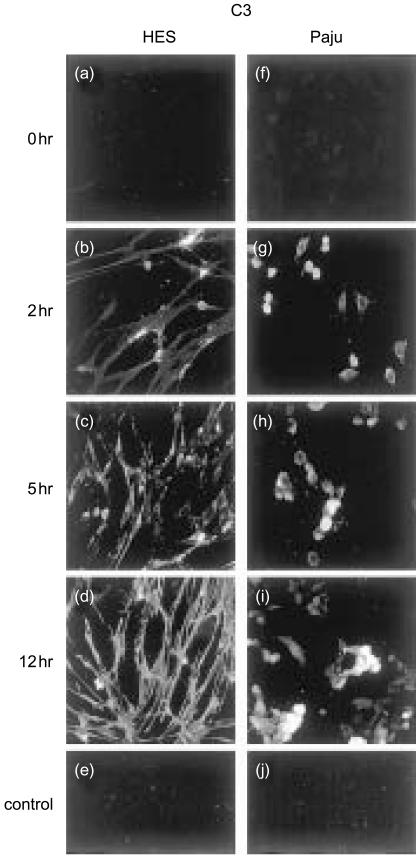
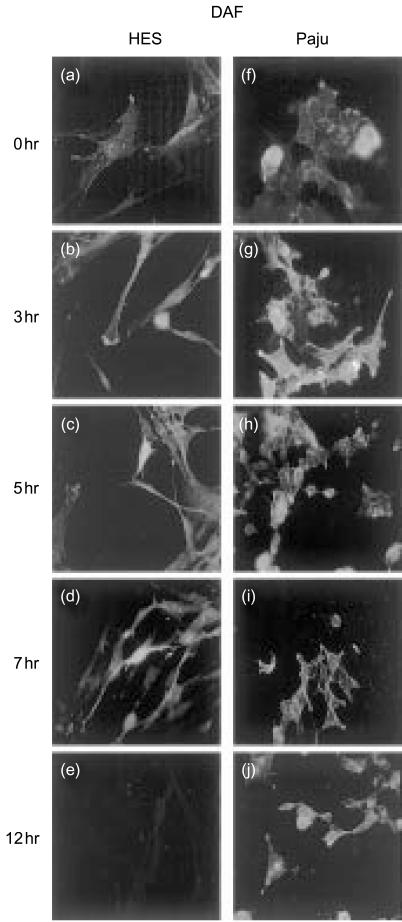
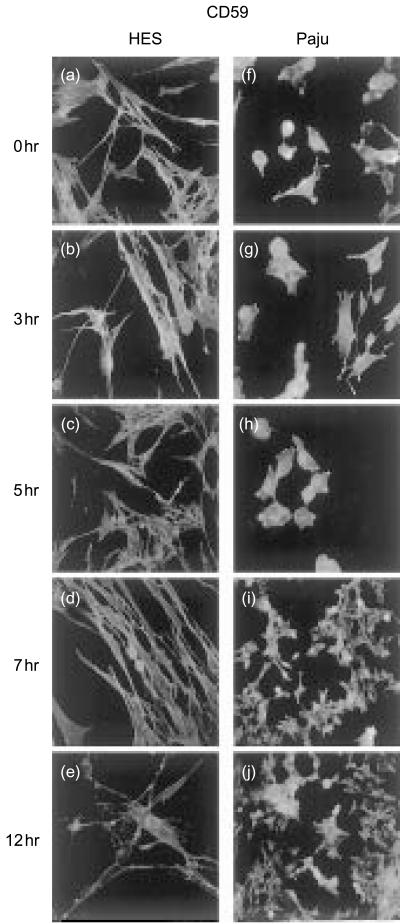
References
-
- Brown EJ, Joiner KA, Frank MM. The role of complement in host resistance to bacteria. Springer Semin Immunopathol. 1983;6:349–60. - PubMed
-
- Joiner KA. Complement evasion by bacteria and parasites. Annu Rev Microbiol. 1988;42:201–30. - PubMed
-
- Grewal AS, Rouse BT. Destruction of virus infected cells by neutrophils and complement. Experientia. 1980;36:352–4. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
