

doi: 10.1101/gad.229402.
The Arabidopsis BODENLOS gene encodes an auxin response protein inhibiting MONOPTEROS-mediated embryo patterning
Affiliations
- PMID: 12101120
- PMCID: PMC186366
- DOI: 10.1101/gad.229402
Item in Clipboard
The Arabidopsis BODENLOS gene encodes an auxin response protein inhibiting MONOPTEROS-mediated embryo patterning
Genes Dev.
.
Abstract
Developmental responses to the plant hormone auxin are thought to be mediated by interacting pairs from two protein families: short-lived inhibitory IAA proteins and ARF transcription factors binding to auxin-response elements. monopteros mutants lacking activating ARF5 and the auxin-insensitive mutant bodenlos fail to initiate the root meristem during early embryogenesis. Here we show that the bodenlos phenotype results from an amino-acid exchange in the conserved degradation domain of IAA12. BODENLOS and MONOPTEROS interact in the yeast two-hybrid assay and the two genes are coexpressed in early embryogenesis, suggesting that BODENLOS inhibits MONOPTEROS action in root meristem initiation.
Figures
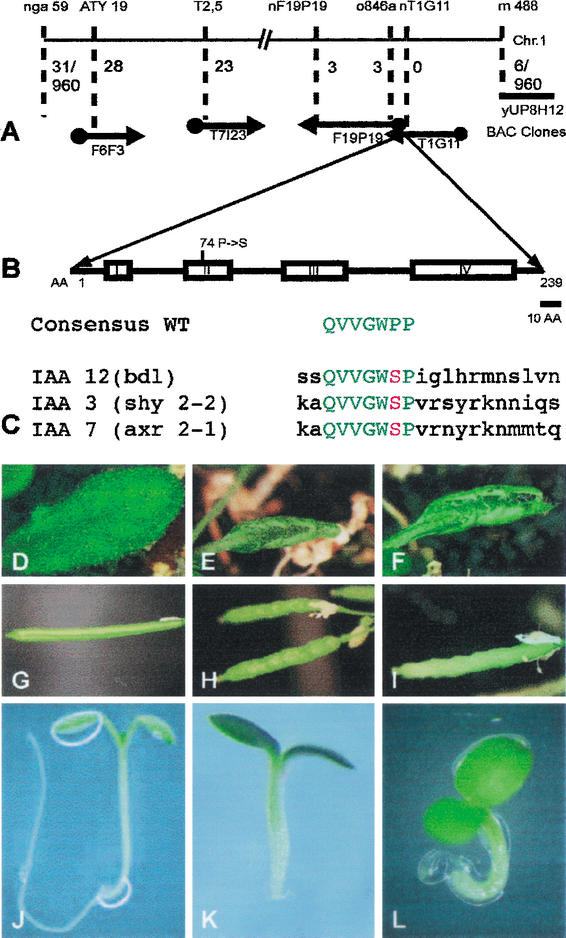
Molecular identification of the BDL gene. (A) Molecular mapping. Molecular markers and BAC clones are shown at the top and bottom, respectively. Numbers indicate recombinants between markers and the BDL gene on BAC clone T1G11 (960 gametes tested). (B) BDL protein domain structure showing the four conserved domains of IAA proteins with the Pro 74 → Ser amino acid exchange in bdl mutant protein. (C) Homologous amino acid exchanges in the conserved domain II of IAA12 (bdl), IAA3 (shy2-2) and IAA7 (axr2-1) mutant proteins. The consensus sequence of wild-type is shown for comparison. (D–L) Phenotypic analysis of transgenic progeny from wild-type plants transformed with the bdl mutant genomic fragment. (D,G,J) Wild-type (BDL/BDL) control; (E,H,K) bdl heterozygous (E,H) and homozygous (K) mutant controls; (F,I,L) transgenic BDL/BDL with one (F,I) or two copies (L) of the bdl transgene. (D–F) Leaves, (G–I) siliques, (J–L) seedlings.
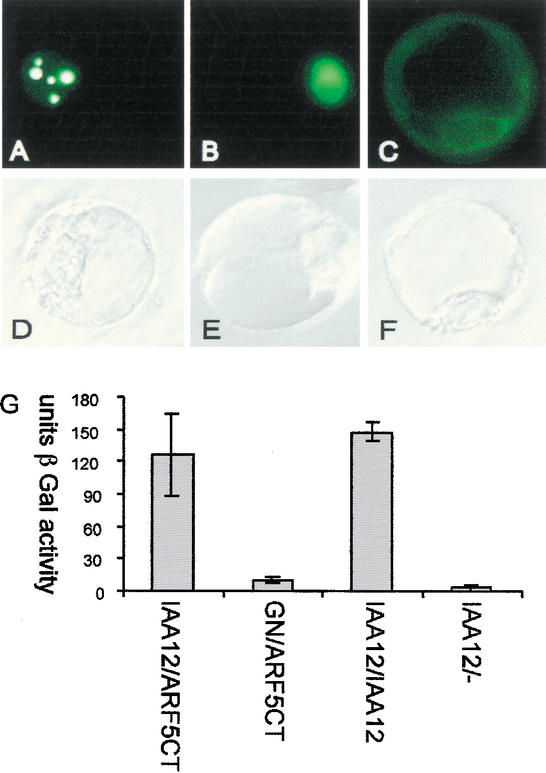
Nuclear localization and interactions of BDL protein. (A–F) Transient expression of 35S∷GFP-BDL in parsley protoplasts. (A–C) GFP fluorescence, (D–F) corresponding bright-field illumination images of (A,D) GFP-BDL, (B,E) GFP with nuclear localization signal, and (C,F) GFP with nuclear export signal. Note strong accumulation of BDL-GFP in patches within the nucleus. (G) Yeast two-hybrid interaction assays. The strength of interaction is shown in activity units of the β-galactosidase reporter (β-Gal). BDL (IAA12) interacts with itself and with the C terminus of MP (ARF5) comprising domains III and IV (ARF5Ct). Negative controls: BDL tested against the empty prey vector (IAA12/−), and GNOM tested against the C terminus of MP (GN/ARF5Ct).
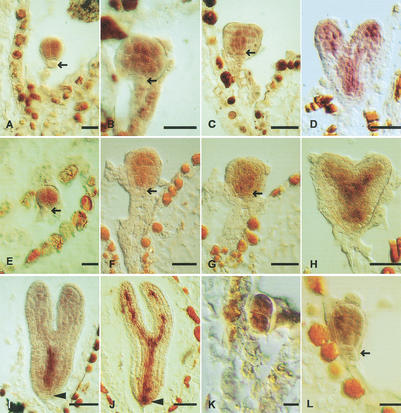
mRNA in situ expression analysis of BDL and MP in developing embryos. (A–D,I) BDL, (E–H,J–L) MP mRNA accumulation in wild-type (E–J) and bdl mutant (K, L) embryos. Developmental stages (Jürgens and Mayer 1994): (A,E) octant; (B,F,L) early-globular; (C,G) globular with lens-shaped cell; (D,H) heart; (I,J) torpedo; (K) four-cell. Arrows mark the hypophysis in (A,B,E,F,L) and the lens-shaped cell in (C,G), arrowheads mark the central root cap in (I,J). Bar, 20 μm.
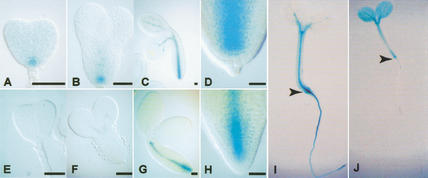
IAA gene promoter-GUS reporter expression in embryos and seedlings. (A–D,I) pBDL∷GUS, (E–H,J) pIAA3∷GUS. Developmental stages (Jürgens and Mayer 1994): (A,E) triangular; (B,F) early-torpedo; (C,G) bent-cotyledon. (D,H) Root end of bent-cotyledon embryos shown at higher magnification. (I,J) Four-day-old seedlings; arrowheads mark the boundary between hypocotyl and root. Bar for A–H, 20 μm.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman GJ, Smith JA, Struhl K. Current protocols in molecular biology. New York, N.Y: Wiley & Sons; 2000.
-
- Berleth T, Jürgens G. The role of the monopteros gene in organising the basal body region of the Arabidopsis embryo. Development. 1993;118:575–587.
-
- Clough SJ, Bent AF. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16:735–743. - PubMed
-
- Fukaki H, Tameda S, Masuda H, Tasaka M. Lateral root formation is blocked by a gain-of-function mutation in the SOLITARY-ROOT/IAA14 gene of Arabidopsis. Plant J. 2002;29:153–168. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases