

Rapid exchange of mammalian topoisomerase II alpha at kinetochores and chromosome arms in mitosis
- PMID: 12105179
- PMCID: PMC2173008
- DOI: 10.1083/jcb.200202053
Rapid exchange of mammalian topoisomerase II alpha at kinetochores and chromosome arms in mitosis
Abstract
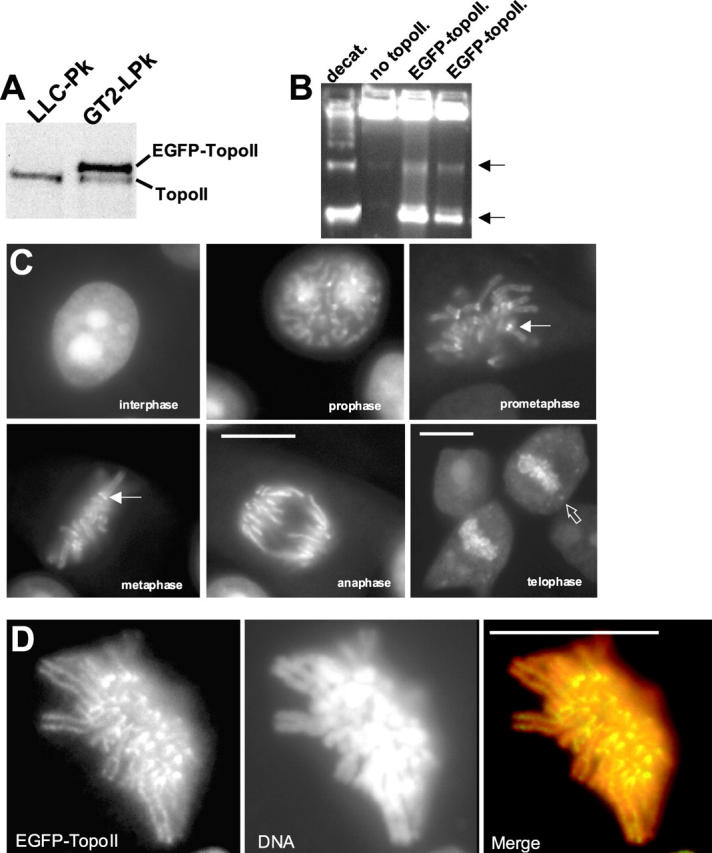
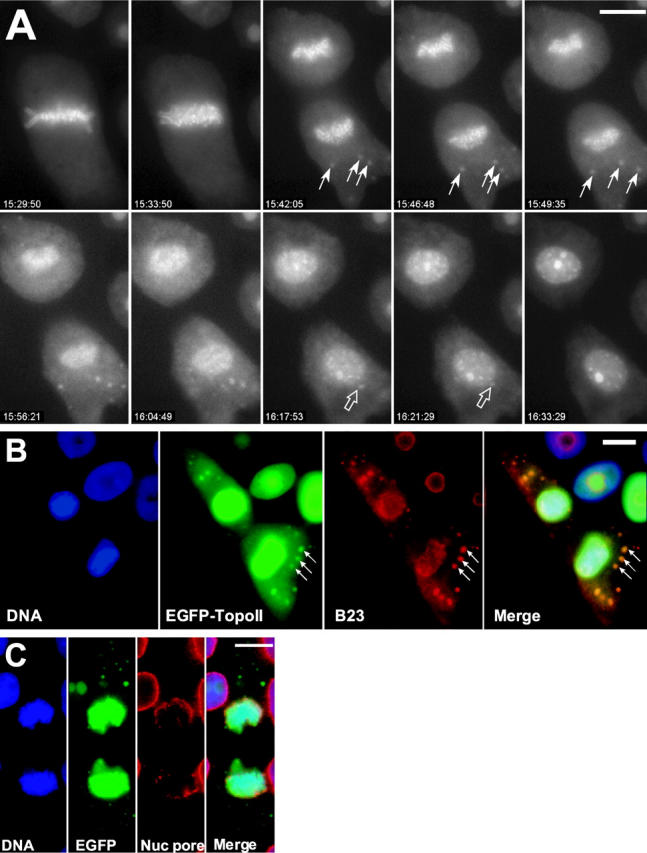
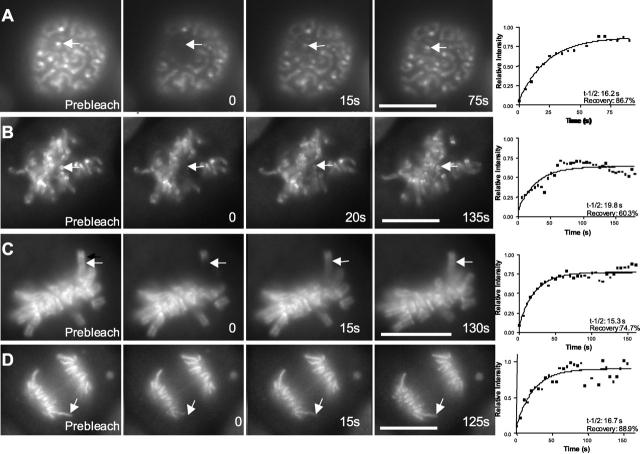
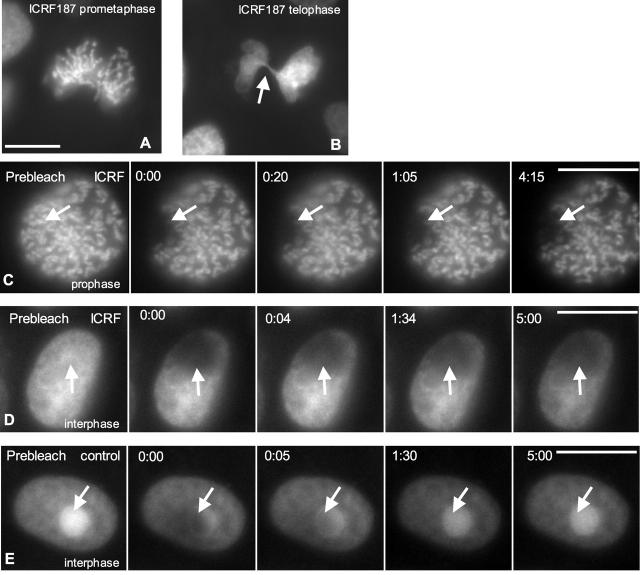
A stable cell line (GT2-LPk) derived from LLC-Pk was created in which endogenous DNA topoisomerase II alpha (topoII alpha) protein was downregulated and replaced by the expression of topoII alpha fused with enhanced green fluorescent protein (EGFP-topoII alpha). The EGFP-topoII alpha faithfully mimicked the distribution of the endogenous protein in both interphase and mitosis. In early stages of mitosis, EGFP-topoII alpha accumulated at kinetochores and in axial lines extending along the chromosome arms. During anaphase, EGFP-topoII alpha diminished at kinetochores and increased in the cytoplasm with a portion accumulating into large circular foci that were mobile and appeared to fuse with the reforming nuclei. These cytoplasmic foci appearing at anaphase were coincident with precursor organelles of the reforming nucleolus called nucleolus-derived foci (NDF). Photobleaching of EGFP-topoII alpha associated with kinetochores and chromosome arms showed that the majority of the protein rapidly exchanges (t1/2 of 16 s). Catalytic activity of topoII alpha was essential for rapid dynamics, as ICRF-187, an inhibitor of topoII alpha, blocked recovery after photobleaching. Although some topoII alpha may be stably associated with chromosomes, these studies indicate that the majority undergoes rapid dynamic exchange. Rapid mobility of topoII alpha in chromosomes may be essential to resolve strain imparted during chromosome condensation and segregation.
Figures
References
-
- Bakshi, R.P., S. Galande, and K. Muniyappa. 2001. Functional and regulatory characteristics of eukaryotic type II DNA topoisomerase. Crit. Rev. Biochem. Mol. Biol. 36:1–37. - PubMed
-
- Barthelmes, H.U., P. Grue, S. Feineis, T. Straub, and F. Boege. 2000. Active DNA topoisomerase IIα is a component of the salt-stable centrosome core. J. Biol. Chem. 275:38823–38830. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
