

Zebrafish trilobite identifies new roles for Strabismus in gastrulation and neuronal movements
- PMID: 12105418
- PMCID: PMC2219916
- DOI: 10.1038/ncb828
Zebrafish trilobite identifies new roles for Strabismus in gastrulation and neuronal movements
Abstract
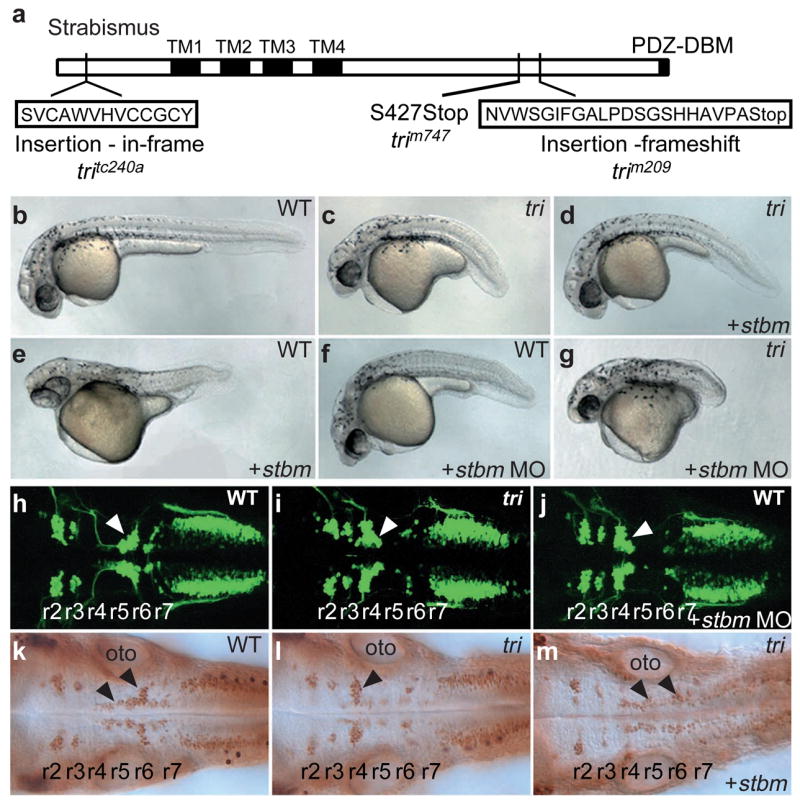
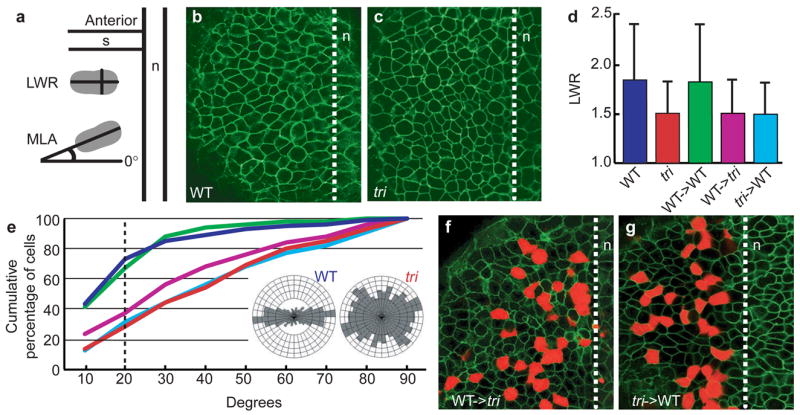
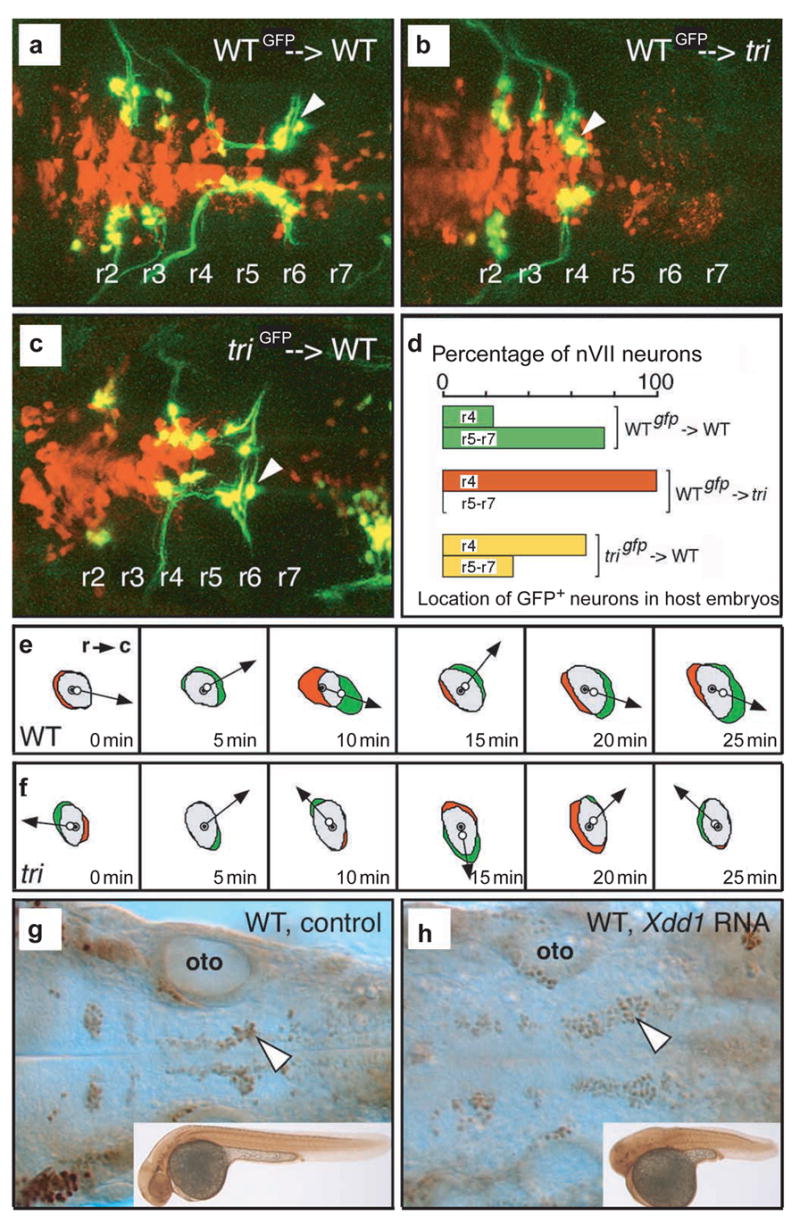
Embryonic morphogenesis is driven by a suite of cell behaviours, including coordinated shape changes, cellular rearrangements and individual cell migrations, whose molecular determinants are largely unknown. In the zebrafish, Dani rerio, trilobite mutant embryos have defects in gastrulation movements and posterior migration of hindbrain neurons. Here, we have used positional cloning to demonstrate that trilobite mutations disrupt the transmembrane protein Strabismus (Stbm)/Van Gogh (Vang), previously associated with planar cell polarity (PCP) in Drosophila melanogaster, and PCP and canonical Wnt/beta-catenin signalling in vertebrates. Our genetic and molecular analyses argue that during gastrulation, trilobite interacts with the PCP pathway without affecting canonical Wnt signalling. Furthermore, trilobite may regulate neuronal migration independently of PCP molecules. We show that trilobite mediates polarization of distinct movement behaviours. During gastrulation convergence and extension movements, trilobite regulates mediolateral cell polarity underlying effective intercalation and directed dorsal migration at increasing velocities. In the hindbrain, trilobite controls effective migration of branchiomotor neurons towards posterior rhombomeres. Mosaic analyses show trilobite functions cell-autonomously and non-autonomously in gastrulae and the hindbrain. We propose Trilobite/Stbm mediates cellular interactions that confer directionality on distinct movements during vertebrate embryogenesis.
Figures

Similar articles
-
Wnt signalling: refocusing on Strabismus.Curr Biol. 2002 Oct 1;12(19):R657-9. doi: 10.1016/s0960-9822(02)01160-0. Curr Biol. 2002. PMID: 12361585
-
Prickle 1 regulates cell movements during gastrulation and neuronal migration in zebrafish.Development. 2003 Sep;130(17):4037-46. doi: 10.1242/dev.00567. Development. 2003. PMID: 12874125
-
Planar cell polarity proteins differentially regulate extracellular matrix organization and assembly during zebrafish gastrulation.Dev Biol. 2013 Nov 1;383(1):39-51. doi: 10.1016/j.ydbio.2013.08.027. Epub 2013 Sep 7. Dev Biol. 2013. PMID: 24021482
-
Core pathway proteins and the molecular basis of planar polarity in the zebrafish gastrula.Semin Cell Dev Biol. 2022 May;125:17-25. doi: 10.1016/j.semcdb.2021.09.015. Epub 2021 Oct 8. Semin Cell Dev Biol. 2022. PMID: 34635444 Review.
-
Cellular and molecular mechanisms of convergence and extension in zebrafish.Curr Top Dev Biol. 2020;136:377-407. doi: 10.1016/bs.ctdb.2019.08.001. Epub 2019 Sep 3. Curr Top Dev Biol. 2020. PMID: 31959296 Free PMC article. Review.
Cited by
-
RNA-seq-based mapping and candidate identification of mutations from forward genetic screens.Genome Res. 2013 Apr;23(4):679-86. doi: 10.1101/gr.147322.112. Epub 2013 Jan 8. Genome Res. 2013. PMID: 23299976 Free PMC article.
-
Regulation of Wnt/PCP signaling through p97/VCP-KBTBD7-mediated Vangl ubiquitination and endoplasmic reticulum-associated degradation.Sci Adv. 2021 May 14;7(20):eabg2099. doi: 10.1126/sciadv.abg2099. Print 2021 May. Sci Adv. 2021. PMID: 33990333 Free PMC article.
-
The planar cell polarity protein VANGL2 coordinates remodeling of the extracellular matrix.Commun Integr Biol. 2012 Jul 1;5(4):325-8. doi: 10.4161/cib.20291. Commun Integr Biol. 2012. PMID: 23060953 Free PMC article.
-
Wnt2 regulates progenitor proliferation in the developing ventral midbrain.J Biol Chem. 2010 Mar 5;285(10):7246-53. doi: 10.1074/jbc.M109.079822. Epub 2009 Dec 16. J Biol Chem. 2010. PMID: 20018874 Free PMC article.
-
Dishevelled genes mediate a conserved mammalian PCP pathway to regulate convergent extension during neurulation.Development. 2006 May;133(9):1767-78. doi: 10.1242/dev.02347. Epub 2006 Mar 29. Development. 2006. PMID: 16571627 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
