

Conformational changes in surface structures of isolated connexin 26 gap junctions
- PMID: 12110573
- PMCID: PMC126111
- DOI: 10.1093/emboj/cdf365
Conformational changes in surface structures of isolated connexin 26 gap junctions
Abstract
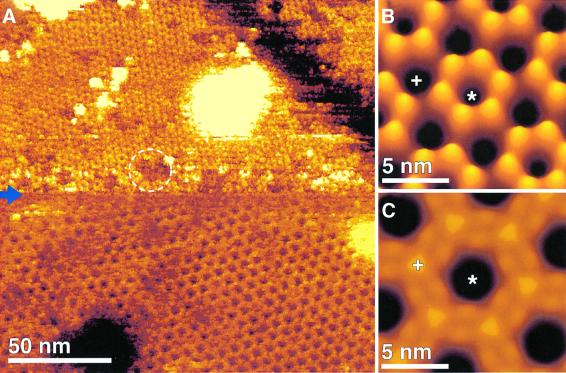
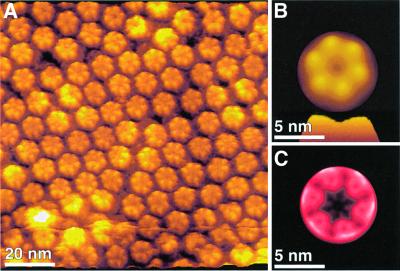
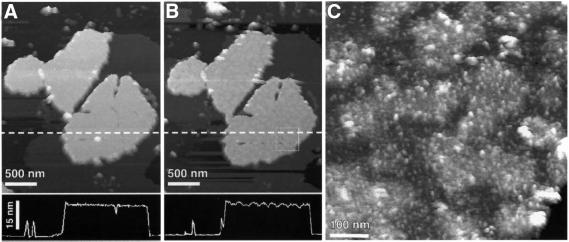
Gap junction channels mediate communication between adjacent cells. Using atomic force microscopy (AFM), we have imaged conformational changes of the cytoplasmic and extracellular surfaces of native connexin 26 gap junction plaques. The cytoplasmic domains of the gap junction surface, imaged at submolecular resolution, form a hexameric pore protruding from the membrane bilayer. Exhibiting an intrinsic flexibility, these cytoplasmic domains, comprising the C-terminal connexin end, reversibly collapse by increasing the forces applied to the AFM stylus. The extracellular connexon surface was imaged after dissection of the gap junction with the AFM stylus. Upon injection of Ca(2+) into the buffer solution, the extracellular channel entrance reduced its diameter from 1.5 to 0.6 nm, a conformational change that is fully reversible and specific among the divalent cations tested. Ca(2+) had a profound effect on the cytoplasmic surface also, inducing the formation of microdomains. Consequently, the plaque height increased by 0.6 nm to 18 nm. This suggests that calcium ions induce conformational changes affecting the structure of both the hemichannels and the intact channels forming cell-cell contacts.
Figures



References
-
- Binnig G., Quate,C.F. and Gerber,C. (1986) Atomic force microscope. Phys. Rev. Lett., 56, 930–933. - PubMed
-
- Czajkowsky D.M. and Shao,Z. (1998) Submolecular resolution of single macromolecules with atomic force microscopy. FEBS Lett., 430, 51–54. - PubMed
-
- Delmar M., Stergiopoulos,K., Homma,N., Calero,G., Morley,G., Ek-Vitorin,J.F. and Taffet,S.M. (2000) A molecular model for the chemical regulation of connexin43 channels: the ‘ball and chain’ hypothesis in gap junctions. In Peracchia,C. (ed.), Molecular Basis of Cell Communication in Health and Disease. Academic Press, San Diego, CA, pp. 223–248.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
