

Ferrichrome induces endosome to plasma membrane cycling of the ferrichrome transporter, Arn1p, in Saccharomyces cerevisiae
- PMID: 12110576
- PMCID: PMC125400
- DOI: 10.1093/emboj/cdf382
Ferrichrome induces endosome to plasma membrane cycling of the ferrichrome transporter, Arn1p, in Saccharomyces cerevisiae
Abstract
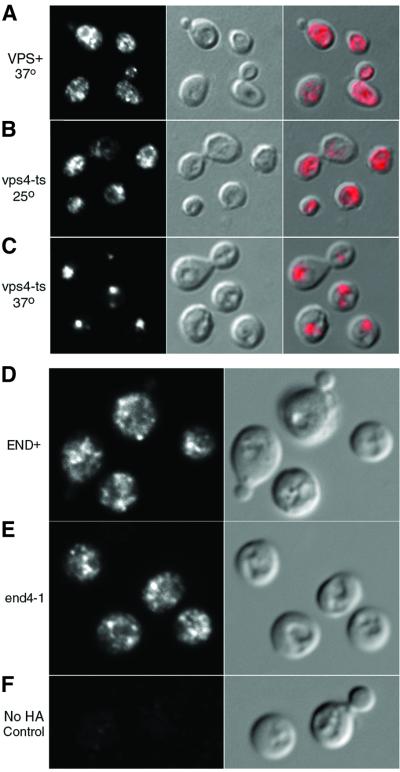
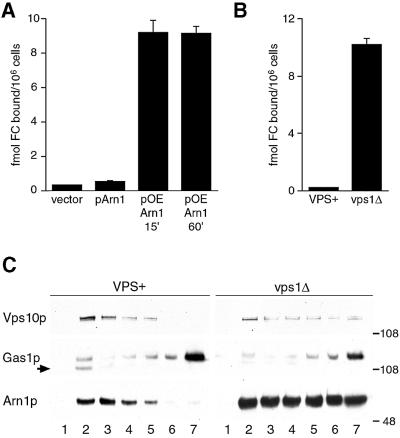
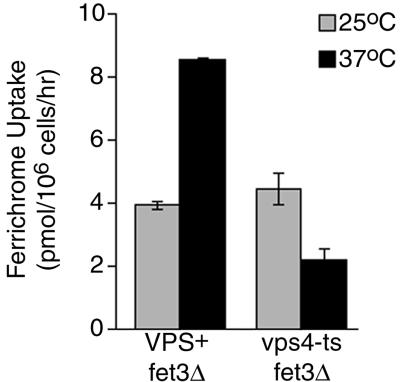
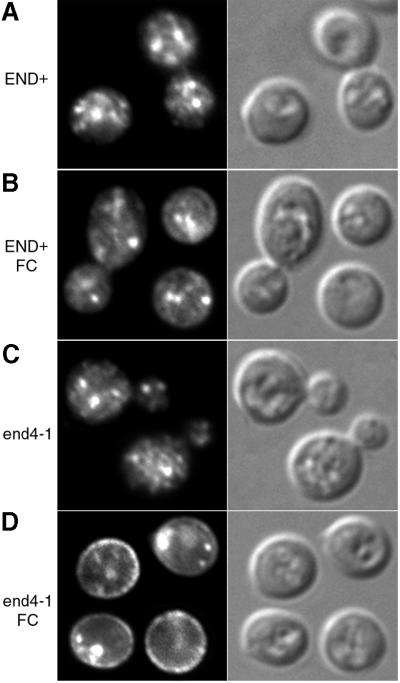
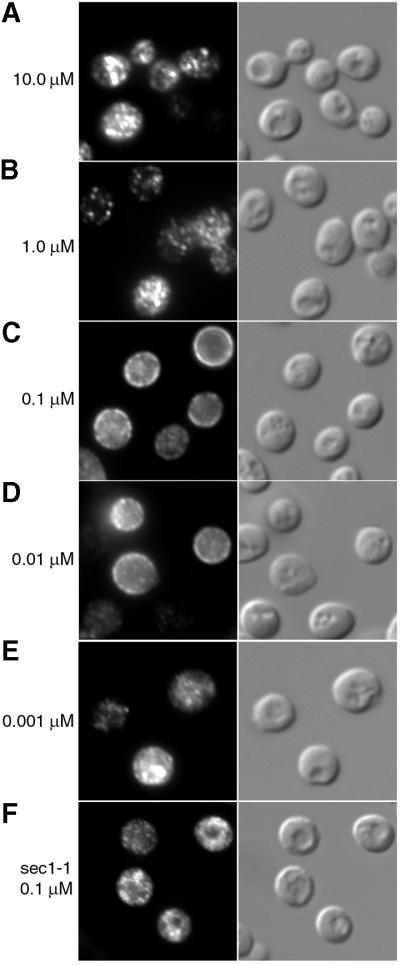
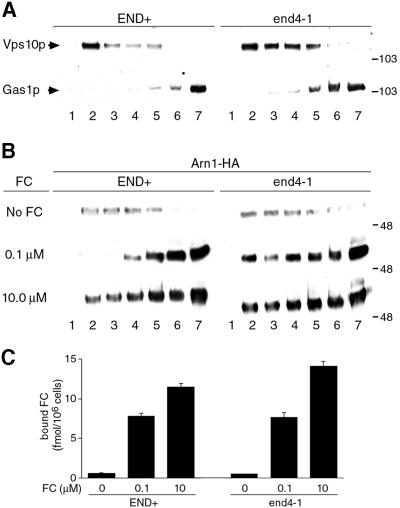
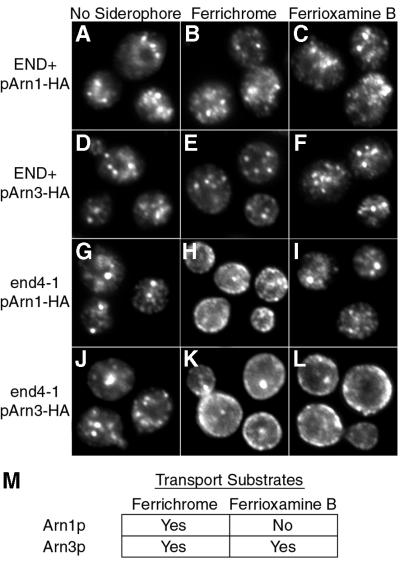
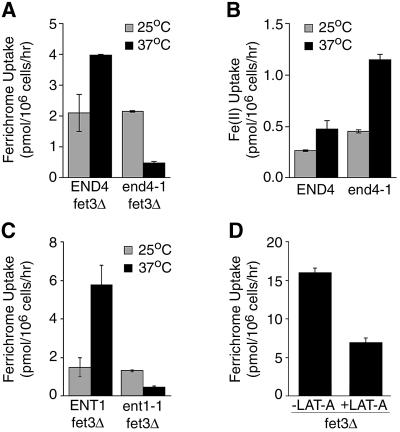
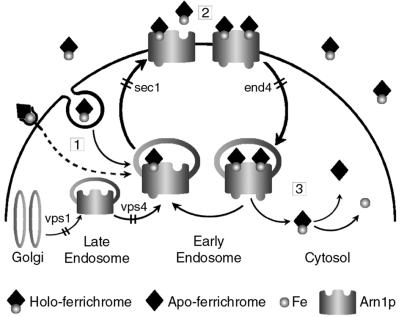
Siderophores are small iron-binding molecules that are synthesized and secreted in the iron-free form by microorganisms. Saccharomyces cerevisiae takes up iron bound to siderophores by two separate systems, one of which requires the ARN family of sidero phore-iron transporters. Arn1p and Arn3p are expressed in endosome-like intracellular vesicles. Here we present evidence that, in the absence of its specific substrate, ferrichrome, Arn1p is sorted directly from the Golgi to the endosomal compartment and does not cycle to the plasma membrane. When cells are exposed to ferrichrome at low concentrations, Arn1p stably relocalizes to the plasma membrane. At higher concentrations of ferrichrome, Arn1p relocalizes to the plasma membrane and rapidly undergoes endocytosis. Plasma membrane localization of Arn1p occurs only in the presence of its specific substrate, and not in the presence of other siderophores. Despite expression of Arn1p on the plasma membrane, mutant strains with defects in endocytosis exhibit reduced uptake of ferrichrome-iron. Thus, siderophores influence the trafficking of the Arn transporters within the cell and this trafficking is important for transporter function.
Figures
Similar articles
-
Siderophore-iron uptake in saccharomyces cerevisiae. Identification of ferrichrome and fusarinine transporters.J Biol Chem. 2000 May 26;275(21):16354-9. doi: 10.1074/jbc.M001456200. J Biol Chem. 2000. PMID: 10748025
-
A receptor domain controls the intracellular sorting of the ferrichrome transporter, ARN1.EMBO J. 2005 Mar 9;24(5):952-62. doi: 10.1038/sj.emboj.7600579. Epub 2005 Feb 17. EMBO J. 2005. PMID: 15719020 Free PMC article.
-
The mechanism of ferrichrome transport through Arn1p and its metabolism in Saccharomyces cerevisiae.Proc Natl Acad Sci U S A. 2003 May 13;100(10):5664-9. doi: 10.1073/pnas.1030323100. Epub 2003 Apr 29. Proc Natl Acad Sci U S A. 2003. PMID: 12721368 Free PMC article.
-
The response to iron deprivation in Saccharomyces cerevisiae: expression of siderophore-based systems of iron uptake.Biochem Soc Trans. 2002 Aug;30(4):698-702. doi: 10.1042/bst0300698. Biochem Soc Trans. 2002. PMID: 12196168 Review.
-
Membrane trafficking in the yeast Saccharomyces cerevisiae model.Int J Mol Sci. 2015 Jan 9;16(1):1509-25. doi: 10.3390/ijms16011509. Int J Mol Sci. 2015. PMID: 25584613 Free PMC article. Review.
Cited by
-
Regulation of cation balance in Saccharomyces cerevisiae.Genetics. 2013 Mar;193(3):677-713. doi: 10.1534/genetics.112.147207. Genetics. 2013. PMID: 23463800 Free PMC article. Review.
-
Expressional control of a cadmium-transporting P1B-type ATPase by a metal sensing degradation signal.J Biol Chem. 2008 Nov 14;283(46):31460-8. doi: 10.1074/jbc.M806054200. Epub 2008 Aug 27. J Biol Chem. 2008. PMID: 18753133 Free PMC article.
-
The Plasma Membrane Protein Nce102 Implicated in Eisosome Formation Rescues a Heme Defect in Mitochondria.J Biol Chem. 2016 Aug 12;291(33):17417-26. doi: 10.1074/jbc.M116.727743. Epub 2016 Jun 17. J Biol Chem. 2016. PMID: 27317660 Free PMC article.
-
New and efficient method using Saccharomyces cerevisiae mutants for identification of siderophores produced by microorganisms.Curr Genet. 2007 Sep;52(3-4):187-90. doi: 10.1007/s00294-007-0145-y. Epub 2007 Jul 31. Curr Genet. 2007. PMID: 17665200
-
Trafficking of siderophore transporters in Saccharomyces cerevisiae and intracellular fate of ferrioxamine B conjugates.Traffic. 2007 Nov;8(11):1601-16. doi: 10.1111/j.1600-0854.2007.00627.x. Epub 2007 Aug 20. Traffic. 2007. PMID: 17714436 Free PMC article.
References
-
- Askwith C.C. and Kaplan,J. (1998) Site-directed mutagenesis of the yeast multicopper oxidase Fet3p. J. Biol. Chem., 273, 22415–22419. - PubMed
-
- Askwith C., Eide,D., Van Ho,A., Bernard,P.S., Li,L., Davis-Kaplan,S., Sipe,D.M. and Kaplan,J. (1994) The FET3 gene of S.cerevisiae encodes a multicopper oxidase required for ferrous iron uptake. Cell, 76, 403–410. - PubMed
-
- Ayscough K. (1998) Use of latrunculin-A, an actin monomer-binding drug. Methods Enzymol., 298, 18–25. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
