

Lesion bypass in yeast cells: Pol eta participates in a multi-DNA polymerase process
- PMID: 12110599
- PMCID: PMC126109
- DOI: 10.1093/emboj/cdf363
Lesion bypass in yeast cells: Pol eta participates in a multi-DNA polymerase process
Abstract
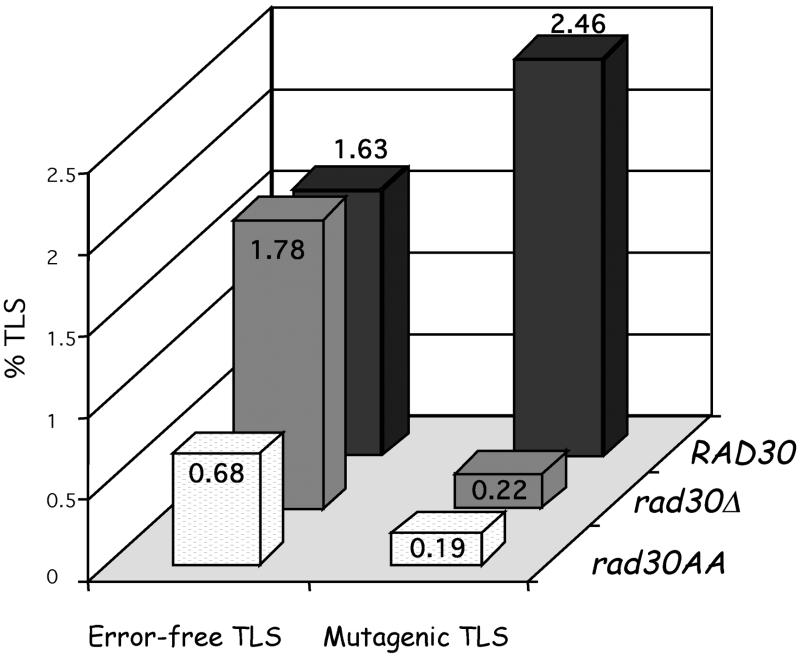
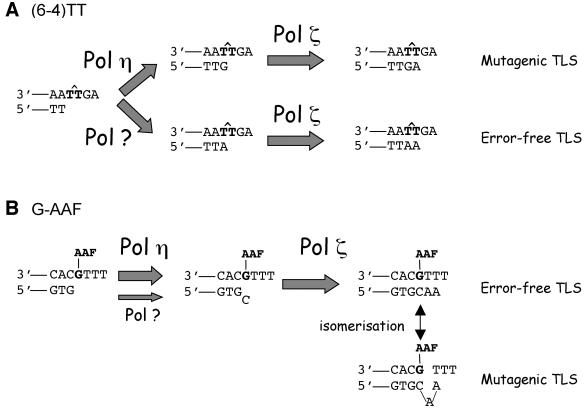
Replication through (6-4)TT and G-AAF lesions was compared in Saccharomyces cerevisiae strains proficient and deficient for the RAD30-encoded DNA polymerase eta (Pol eta). In the RAD30 strain, the (6-4)TT lesion is replicated both inaccurately and accurately 60 and 40% of the time, respectively. Surprisingly, in a rad30 Delta strain, the level of mutagenic bypass is essentially suppressed, while error-free bypass remains unchanged. Therefore, Pol eta is responsible for mutagenic replication through the (6-4)TT photoproduct, while another polymerase mediates its error-free bypass. Deletion of the RAD30 gene also reduces the levels of both accurate and inaccurate bypass of AAF lesions within two different sequence contexts up to 8-fold. These data show that, in contrast to the accurate bypass by Pol eta of TT cyclobutane dimers, it is responsible for the mutagenic bypass of other lesions. In conclusion, this paper shows that, in yeast, translesion synthesis involves the combined action of several polymerases.
Figures
Similar articles
-
Specificity of DNA lesion bypass by the yeast DNA polymerase eta.J Biol Chem. 2000 Mar 17;275(11):8233-9. doi: 10.1074/jbc.275.11.8233. J Biol Chem. 2000. PMID: 10713149
-
Unlocking the steric gate of DNA polymerase η leads to increased genomic instability in Saccharomyces cerevisiae.DNA Repair (Amst). 2015 Nov;35:1-12. doi: 10.1016/j.dnarep.2015.07.002. Epub 2015 Aug 7. DNA Repair (Amst). 2015. PMID: 26340535 Free PMC article.
-
Mutator effects of overproducing DNA polymerase eta (Rad30) and its catalytically inactive variant in yeast.Mutat Res. 2001 Jul 1;478(1-2):129-39. doi: 10.1016/s0027-5107(01)00131-2. Mutat Res. 2001. PMID: 11406177
-
DNA polymerase eta and chemotherapeutic agents.Antioxid Redox Signal. 2011 Jun 15;14(12):2521-9. doi: 10.1089/ars.2010.3673. Epub 2011 Mar 18. Antioxid Redox Signal. 2011. PMID: 21050139 Free PMC article. Review.
-
The accurate bypass of pyrimidine dimers by DNA polymerase eta contributes to ultraviolet-induced mutagenesis.Mutat Res. 2024 Jan-Jun;828:111840. doi: 10.1016/j.mrfmmm.2023.111840. Epub 2023 Nov 7. Mutat Res. 2024. PMID: 37984186 Review.
Cited by
-
Error-free replicative bypass of thymine glycol by the combined action of DNA polymerases kappa and zeta in human cells.Proc Natl Acad Sci U S A. 2010 Aug 10;107(32):14116-21. doi: 10.1073/pnas.1007795107. Epub 2010 Jul 26. Proc Natl Acad Sci U S A. 2010. PMID: 20660785 Free PMC article.
-
Error-free replicative bypass of (6-4) photoproducts by DNA polymerase zeta in mouse and human cells.Genes Dev. 2010 Jan 15;24(2):123-8. doi: 10.1101/gad.1872810. Genes Dev. 2010. PMID: 20080950 Free PMC article.
-
Canonical and Non-Canonical Roles of Human DNA Polymerase η.Genes (Basel). 2024 Sep 27;15(10):1271. doi: 10.3390/genes15101271. Genes (Basel). 2024. PMID: 39457395 Free PMC article. Review.
-
Mms2-Ubc13-dependent and -independent roles of Rad5 ubiquitin ligase in postreplication repair and translesion DNA synthesis in Saccharomyces cerevisiae.Mol Cell Biol. 2006 Oct;26(20):7783-90. doi: 10.1128/MCB.01260-06. Epub 2006 Aug 14. Mol Cell Biol. 2006. PMID: 16908531 Free PMC article.
-
Damage mapping techniques and the light they have shed on canonical and atypical UV photoproducts.Front Genet. 2023 Jan 10;13:1102593. doi: 10.3389/fgene.2022.1102593. eCollection 2022. Front Genet. 2023. PMID: 36704334 Free PMC article. Review.
References
-
- Baynton K. and Fuchs,R.P.P. (2000) Lesions in DNA: hurdles for polymerases. Trends Biochem. Sci., 25, 74–79. - PubMed
-
- Baynton K., Bresson-Roy,A. and Fuchs,R.P. (1999) Distinct roles for Rev1p and Rev7p during translesion synthesis in Saccharomyces cerevisiae. Mol. Microbiol., 34, 124–133. - PubMed
-
- Becherel O.J. and Fuchs,R.P. (1999) SOS mutagenesis results from up-regulation of translesion synthesis. J. Mol. Biol., 294, 299–306. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
