

Accessory factors determine the order of strand exchange in Xer recombination at psi
- PMID: 12110600
- PMCID: PMC126124
- DOI: 10.1093/emboj/cdf379
Accessory factors determine the order of strand exchange in Xer recombination at psi
Abstract
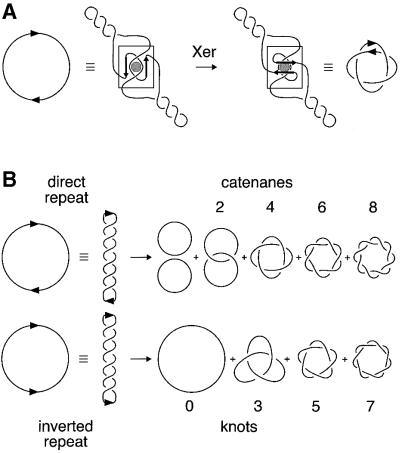
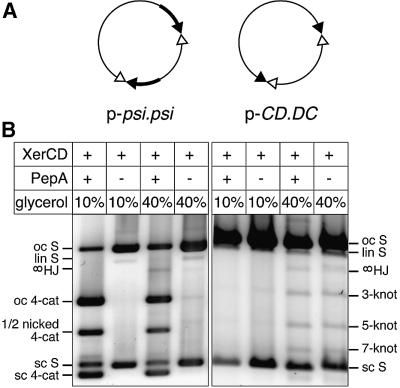
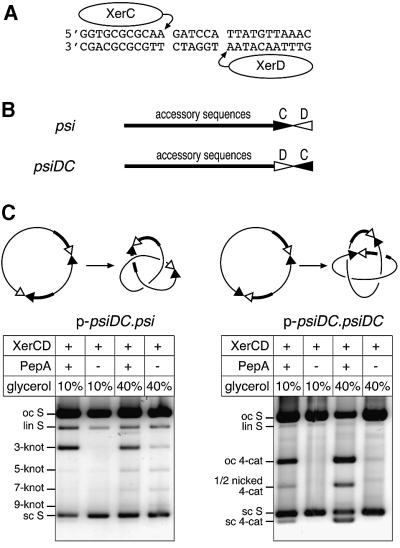
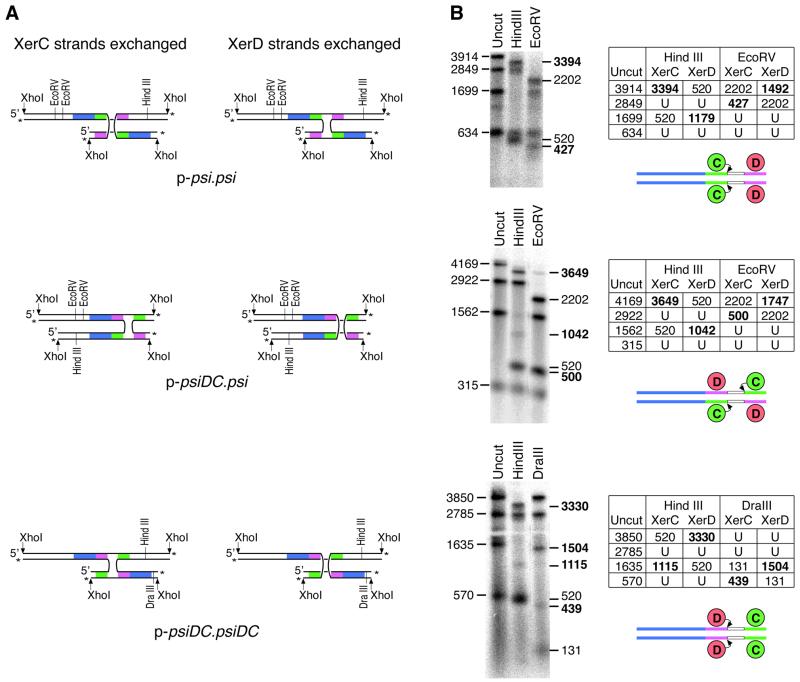
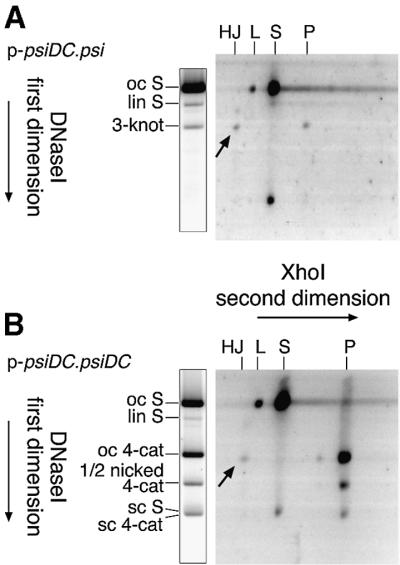
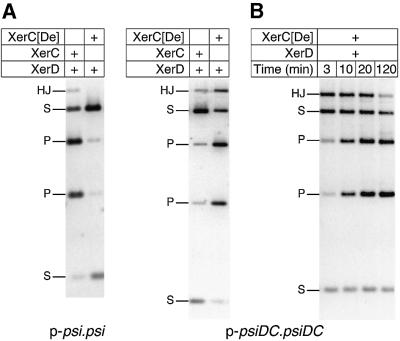
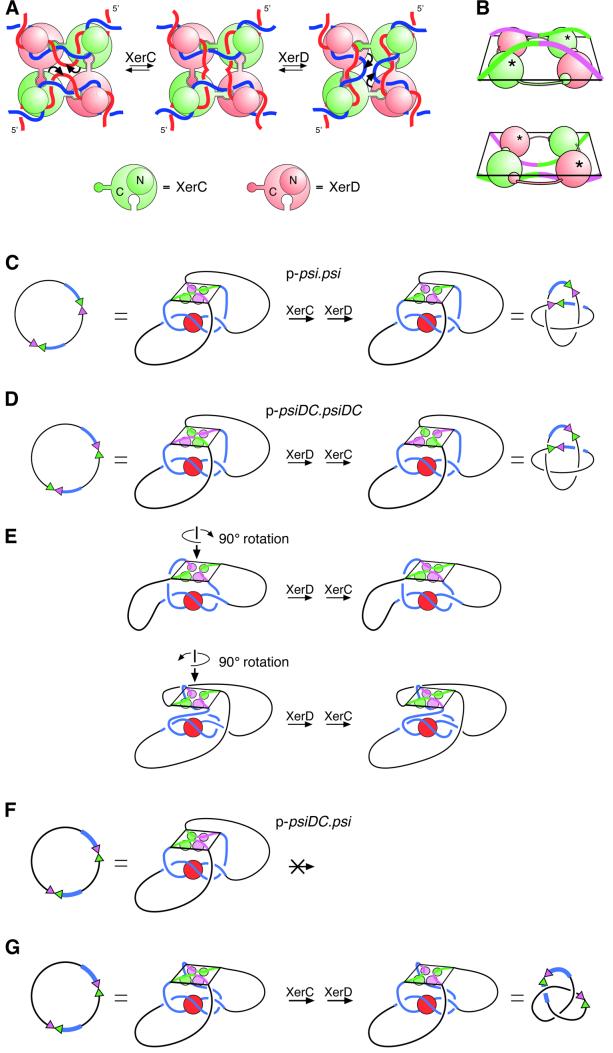
Xer site-specific recombination in Escherichia coli converts plasmid multimers to monomers, thereby ensuring their correct segregation at cell division. Xer recombination at the psi site of plasmid pSC101 is preferentially intramolecular, giving products of a single topology. This intramolecular selectivity is imposed by accessory proteins, which bind at psi accessory sequences and activate Xer recombination at the psi core. Strand exchange proceeds sequentially within the psi core; XerC first exchanges top strands to produce Holliday junctions, then XerD exchanges bottom strands to give final products. In this study, recombination was analysed at sites in which the psi core was inverted with respect to the accessory sequences. A plasmid containing two inverted-core psi sites recombined with a reversed order of strand exchange, but with unchanged product topology. Thus the architecture of the synapse, formed by accessory proteins binding to accessory sequences, determines the order of strand exchange at psi. This finding has important implications for the way in which accessory proteins interact with the recombinases.
Figures
References
-
- Arciszewska L.K., Baker,R.A., Hallet,B. and Sherratt,D.J. (2000) Coordinated control of XerC and XerD catalytic activities during Holliday junction resolution. J. Mol. Biol., 299, 391–403. - PubMed
-
- Aussel L., Barre,F.-X., Aroyo,M., Stasiak,A., Stasiak,A.Z. and Sherratt,D.J. (2002) FtsK is a DNA motor protein that activates chromosome dimer resolution by switching the catalytic state of XerC and XerD recombinases. Cell, 108, 195–205. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
