

Phosphite, an analog of phosphate, suppresses the coordinated expression of genes under phosphate starvation
- PMID: 12114577
- PMCID: PMC166517
- DOI: 10.1104/pp.010835
Phosphite, an analog of phosphate, suppresses the coordinated expression of genes under phosphate starvation
Abstract
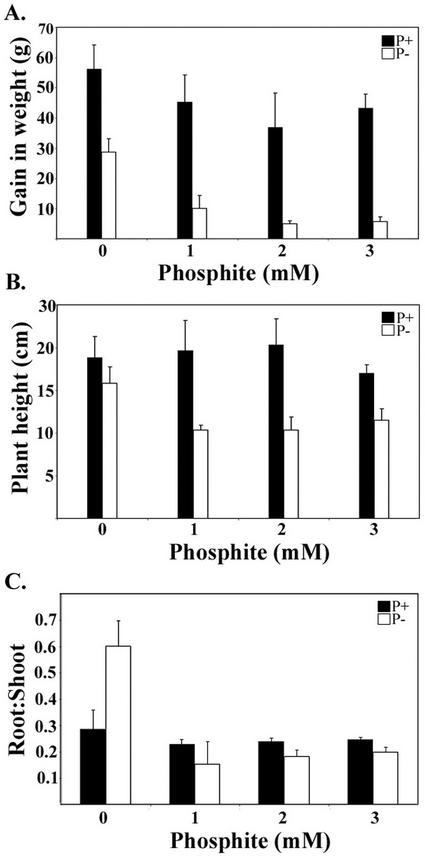
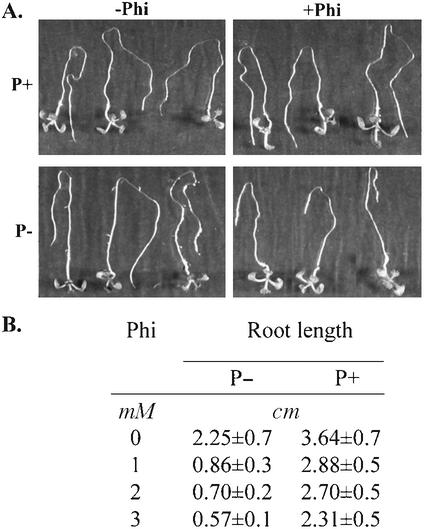
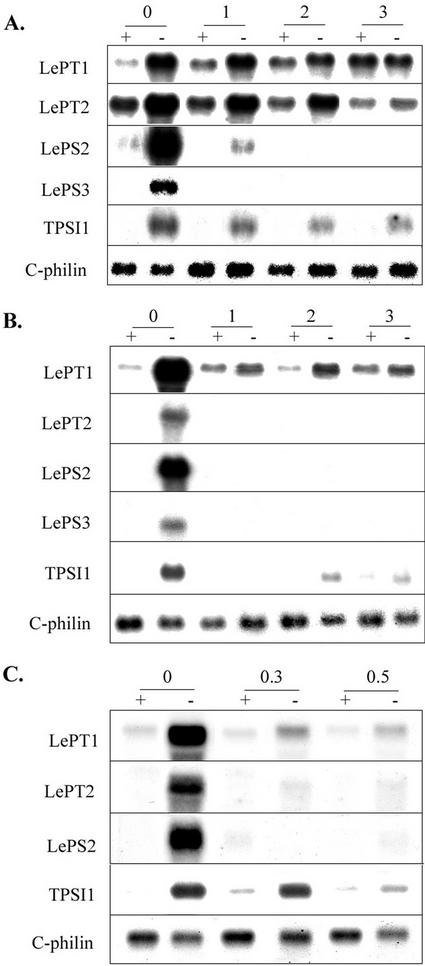
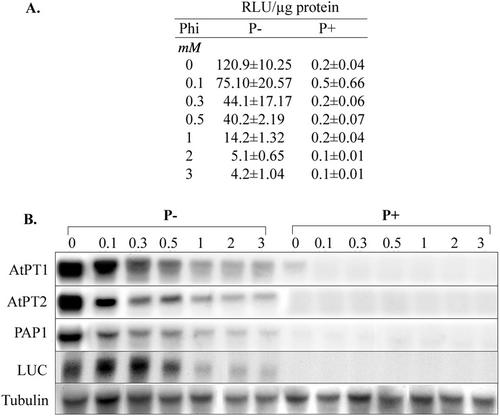
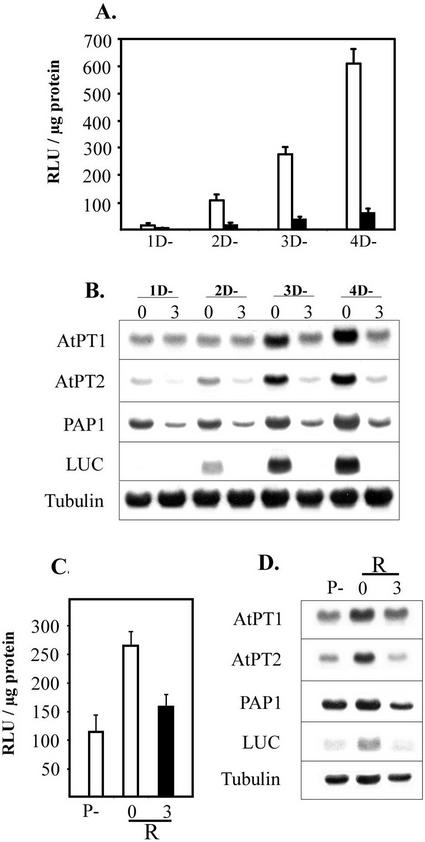
Phosphate (Pi) and its analog phosphite (Phi) are acquired by plants via Pi transporters. Although the uptake and mobility of Phi and Pi are similar, there is no evidence suggesting that plants can utilize Phi as a sole source of phosphorus. Phi is also known to interfere with many of the Pi starvation responses in plants and yeast (Saccharomyces cerevisiae). In this study, effects of Phi on plant growth and coordinated expression of genes induced by Pi starvation were analyzed. Phi suppressed many of the Pi starvation responses that are commonly observed in plants. Enhanced root growth and root to shoot ratio, a hallmark of Pi stress response, was strongly inhibited by Phi. The negative effects of Phi were not obvious in plants supplemented with Pi. The expression of Pi starvation-induced genes such as LePT1, LePT2, AtPT1, and AtPT2 (high-affinity Pi transporters); LePS2 (a novel acid phosphatase); LePS3 and TPSI1 (novel genes); and PAP1 (purple acid phosphatase) was suppressed by Phi in plants and cell cultures. Expression of luciferase reporter gene driven by the Pi starvation-induced AtPT2 promoter was also suppressed by Phi. These analyses showed that suppression of Pi starvation-induced genes is an early response to addition of Phi. These data also provide evidence that Phi interferes with gene expression at the level of transcription. Synchronized suppression of multiple Pi starvation-induced genes by Phi points to its action on the early molecular events, probably signal transduction, in Pi starvation response.
Figures
References
-
- Baldwin JC, Karthikeyan AS, Raghothama KG. Annual American Society of Plant Physiologists Meeting, July 24–28, Baltimore. Rockville, MD: American Society of Plant Physiologists; 1999. LePS3, a novel phosphate starvation induced gene in tomato (abstract no. 937) p. 190.
-
- Barchietto T, Saindrenan P, Bompeix G. Characterization of phosphonate uptake in two Phytophthora spp. and its inhibition by phosphate. Arch Microbiol. 1989;151:54–58.
-
- Barchietto T, Saindrenan P, Bompeix G. Physiological responses of Phytophthora citrophthora to a sub-inhibitory concentration of phosphonate. Pestic Biochem Physiol. 1992;42:151–166.
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
