

Patterns of epithelial cell invasion by different species of the Burkholderia cepacia complex in well-differentiated human airway epithelia
- PMID: 12117967
- PMCID: PMC128168
- DOI: 10.1128/IAI.70.8.4547-4555.2002
Patterns of epithelial cell invasion by different species of the Burkholderia cepacia complex in well-differentiated human airway epithelia
Abstract
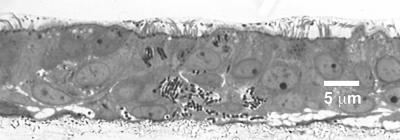
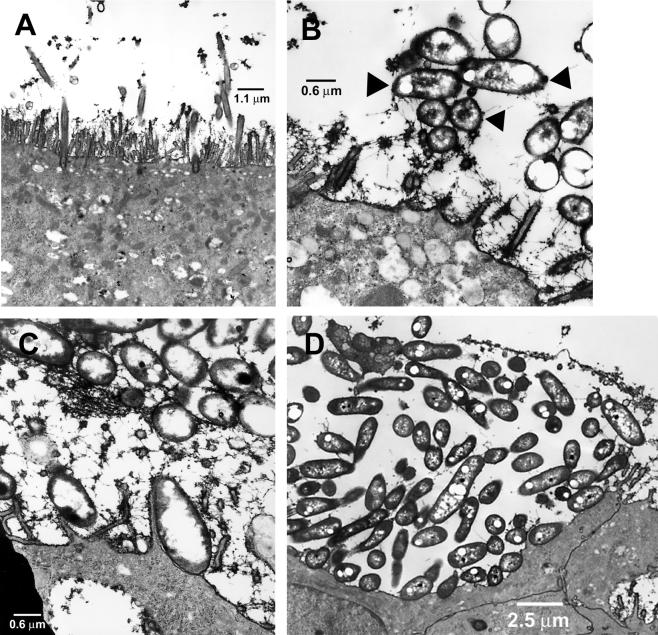
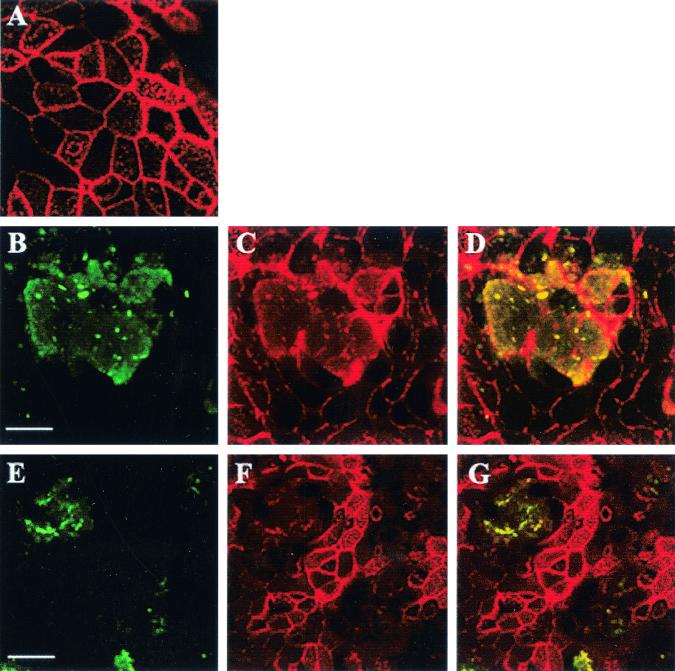
Burkholderia cepacia has emerged as a serious respiratory pathogen in cystic fibrosis (CF) patients. The clinical course of B. cepacia infections is variable, but approximately 20% of patients eventually succumb to the cepacia syndrome, which is characterized as a fatal necrotizing pneumonia with bacteremia. The mechanisms that permit B. cepacia to cause bacteremia are not yet known but probably involve sequential penetration of airway barriers. This study evaluated the abilities of different species of the B. cepacia complex, including a strain from the ET12 lineage (BC-7, genomovar III, cblA(+)), which is associated with most cepacia syndrome fatalities among CF populations, a genomovar IV strain (HI2258), and a genomovar II strain (J-1) to penetrate polarized, well-differentiated human airway epithelial cell cultures. As revealed by light and electron microscopy, all three B. cepacia strains tested circumvented the mechanical barriers of mucus and ciliary transport to penetrate the airway epithelium but they used different routes. The BC-7 strain (genomovar III) formed biofilms in close proximity to the apical cell surface, followed by invasion and destruction of epithelial cells. This process involved disruption of the glycocalyx and rearrangements of the actin cytoskeleton. The HI2258 strain (genomovar IV) did not form biofilms, and the majority of bacteria that penetrated the epithelium were located between epithelial cells, suggesting paracytosis. Strain J-1 penetrated the epithelium both by cell destruction and paracytosis. These studies suggest that the distinct invasion pathways employed by B. cepacia may account for differences in virulence between B. cepacia genomovars.
Figures


Similar articles
-
Disproportionate distribution of Burkholderia cepacia complex species and transmissibility markers in cystic fibrosis.Am J Respir Crit Care Med. 2001 Jul 1;164(1):92-6. doi: 10.1164/ajrccm.164.1.2011153. Am J Respir Crit Care Med. 2001. PMID: 11435245
-
Infection with Burkholderia cepacia complex genomovars in patients with cystic fibrosis: virulent transmissible strains of genomovar III can replace Burkholderia multivorans.Clin Infect Dis. 2001 Nov 1;33(9):1469-75. doi: 10.1086/322684. Epub 2001 Oct 4. Clin Infect Dis. 2001. PMID: 11588691
-
Lack of cable pili expression by cblA-containing Burkholderia cepacia complex.Microbiology (Reading). 2002 Nov;148(Pt 11):3477-3484. doi: 10.1099/00221287-148-11-3477. Microbiology (Reading). 2002. PMID: 12427939
-
Burkholderia cepacia infection and lung transplantation.Semin Respir Infect. 2002 Dec;17(4):284-90. doi: 10.1053/srin.2002.36443. Semin Respir Infect. 2002. PMID: 12497545 Review.
-
Intracellular survival and innate immune evasion of Burkholderia cepacia: Improved understanding of quorum sensing-controlled virulence factors, biofilm, and inhibitors.Microbiol Immunol. 2020 Feb;64(2):87-98. doi: 10.1111/1348-0421.12762. Epub 2020 Jan 10. Microbiol Immunol. 2020. PMID: 31769530 Review.
Cited by
-
Virulence traits associated with Burkholderia cenocepacia ST856 epidemic strain isolated from cystic fibrosis patients.Antimicrob Resist Infect Control. 2017 Jun 6;6:57. doi: 10.1186/s13756-017-0215-y. eCollection 2017. Antimicrob Resist Infect Control. 2017. PMID: 28593045 Free PMC article.
-
Burkholderia cepacia Complex Vaccines: Where Do We Go from here?Vaccines (Basel). 2016 Apr 15;4(2):10. doi: 10.3390/vaccines4020010. Vaccines (Basel). 2016. PMID: 27092530 Free PMC article. Review.
-
The tyrosine kinase BceF and the phosphotyrosine phosphatase BceD of Burkholderia contaminans are required for efficient invasion and epithelial disruption of a cystic fibrosis lung epithelial cell line.Infect Immun. 2015 Feb;83(2):812-21. doi: 10.1128/IAI.02713-14. Epub 2014 Dec 8. Infect Immun. 2015. PMID: 25486990 Free PMC article.
-
Invited review: human air-liquid-interface organotypic airway tissue models derived from primary tracheobronchial epithelial cells-overview and perspectives.In Vitro Cell Dev Biol Anim. 2021 Feb;57(2):104-132. doi: 10.1007/s11626-020-00517-7. Epub 2020 Nov 11. In Vitro Cell Dev Biol Anim. 2021. PMID: 33175307 Free PMC article. Review.
-
Best Practices in the Development and Use of Experimental Models of Bacterial Pneumonia: An Official American Thoracic Society Workshop Report.Am J Respir Cell Mol Biol. 2025 Aug;73(2):178-199. doi: 10.1165/rcmb.2025-0322ST. Am J Respir Cell Mol Biol. 2025. PMID: 40748059 Free PMC article.
References
-
- Bernacki, S. H., A. L. Nelson, L. Abdullah, J. K. Sheehan, A. Harris, C. W. Davis, and S. H. Randell. 1999. Mucin gene expression during differentiation of human airway epithelia in vitro. Am. J. Respir. Cell Mol. Biol. 20:595-604. - PubMed
-
- Buhler,T., S. Ballestero, M. Desai, and M. R. Brown. 1998. Generation of a reproducible nutrient-depleted biofilm of Escherichia coli and Burkholderia cepacia. J. Appl. Microbiol. 85:457-462. - PubMed
-
- Costerton, J. W., P. S. Stewart, and E. P. Greenberg. 1999. Bacterial biofilms: a common cause of persistent infections. Science 284:1318-1322. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
